











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
The sustainability of beef cattle production systems in relation to concerns about climate change and the quality of the services provided to society has become a fundamental issue for public debate. However, beef cattle production systems on pasture differ in terms of the use of resources, degree of intensification, cultural roles, among others. At least 50% of Latin America and the Caribbean is either too dry, too wet, too steep, too shallow, too infertile, or too fragile to sustain cultivation or to support forests. Analyzing sustainability at the farm level for gaining an understanding of the reproducibility of pastoral systems should involve the management of animals and grazing resources, their economic performance, and environmental implications.
Grasslands are too often neglected in national land resource inventories and are given pejoratives as "uncultivated, idle or underutilized land," but, in Venezuela, savannahs and grasslands occupy 57% of the total agricultural land (cropland and forest included). If it were possible to measure the energy consumed by the grazing herd in the equivalent of grain production, the result would be staggering. In fact, 13+ million heads of cattle consumed in 2007, only for maintenance, the equivalent of 12,3 million MT of yellow corn: 5,1 times more than the record corn harvest of 2007 (INE, 2007). It is about the proper use of this resource what matters.
On the efficiency and productivity of Beef Cattle Production Systems on Pasture significant progress could be made in terms of adoption of improved management procedures: Individual identification of the animal to permit proper record-keeping; Generalized use of the breeding season, allowing better synchronization of nutritive resources and herd requirements (Arriaga, 2010); The systematic weaning and the consequent segregation of the herd by sex, age, and production status will allow the use of a limited supplement of younger animals; Development of applied techniques of forage conservation where tame pasture grass is available, as well as range improvement and soil conservation practices, (Tejos, 2014); General improvement of communication and transportation; more extensive use of mineral supplementation (Depablos, Ordóñez, Godoy, & Chicco, 2009), vaccination, dipping and a higher level of sanitary control (Pierre & Camaripano, 2009; Camaripano, Reina, & Plasse, 2011). All the above-mentioned practices are currently being used in experimental stations and elite herds (Verde, Medina, & Borges, 2007; Depablos, Ojeda, Martínez, & Colmenares, 2010). Their use has accounted for an almost doubling of the production over the commercial producer's normal level (Plasse, 2000).
Dr. Gordon E. Dickerson was a visionary and productive scientist whose many scientific contributions built the scaffolding of the systems approach to the genetic improvement of the economic efficiency of beef production (Tess & Davis, 2002). G. Dickerson's contributions to understanding heterosis and epistasis rank among his most important works (Dickerson, 1969; Dickerson, 1973). These studies demonstrated that economic efficiency was most improved in systems that exploited both individual and maternal heterosis.
Hybrid vigor or heterosis is the superiority of the crosses over the average of the parent breeds. Bunning, Wall, Chagunda, Banos, & Simm, (2019) performed a meta-analysis of heterosis in tropical cattle, concluding: Heterosis was found to be beneficial for a range of economically important traits, including those related to fitness such as fertility and longevity, which are particularly important in low input systems prevalent in the tropics. The greatest heterosis was expressed in crosses of breeds adapted to different environments; they allow the combination of complementary production and fitness traits, meaning that there is great potential to utilize heterosis to increase profitability.
To assess the magnitude of heterosis in each environment, it is necessary to compare F1 crosses with the average of both pure parents (Plasse, 2000). Some estimates of the effects of heterosis in Criollo - Bos indicus crosses have been published in Venezuela. Ordóñez et al. (1974) reported for age (-72.0 days) and weight (35.2 kg) at puberty of heifers, while Plasse (1983) summarized the available average values (variation range between experiments in parentheses) for the different characteristics: pregnancy percentage, 14 % (9-16 %); weaning weight, 11 % (9-13 %); post-weaning weight 16 % (12-19 %). In crosses among Zebu and Bos taurus breeds other than Criollo, due to lack of adaptation in pure form only Ordóñez (1985) estimated heterosis for birth weight (-0.5 kg), weaning weight (3.0 kg) and 18 months weight (16.1 kg) in crosses Brahman X Charolais to conclude that: (1) The low adaptability of the Charolais cow in pure breeding prevented the expression of the crossbred genotypes in pre-weaning traits, while the post-weaning poor adaptation of the Charolais progeny, increased the heterosis in post-weaning gain; (2) Heterosis is not a parameter determined by the genotypes that intervene in the crossing, but its expression is conditioned by the environment; (3) The equations applied to estimate performance using mean breed effects and expected heterosis might be inadequate when environmental limitations or lack of adaptability prevent the expression of genetic potentials; (4) Although the superiority of the crosses over the average of the parent breeds is explained in genetic terms as the increased heterozygosity in the hybrid, the phenotypic expression of the superiority observed in hybrids relative to their parents, for some traits, is the result of production potentials that are only reached in an adequate environment, both must be considered for the definition of genetic programs.
When developing breeding systems applicable to the tropics, some conditions must be accomplished. To be useful, the system must: (1) Allow female replacements to be generated throughout the herd; (2) Effectively exploit heterosis; (3) Do not interfere with selection for additive traits; (4) Both females and males must be fully adapted to the conditions under which they will have to work.
Rotational crossbreeding seems to be the only mating system that fulfills these conditions. In a previous paper, (Ordóñez, 1975) pointed out the shortcomings of rotation: (1) Complicated handling (more than half the herds keep one or at most two bulls) and (2) Divergent genetic composition between successive generations. This work aims to evaluate and compare the advantages of alternative mating systems proposed: (A) Composites or the development of new breeds;(B) Modified Absorption system where the bulls used on private holdings are F1.
Methodology
This work was based on bibliographic research about Mating Systems in beef cattle production systems and their application in the American Tropics, with emphasis on Venezuela. It reviews the theoretical basis of hybrid vigor or heterosis (Cartwright, 1970; Dickerson, 1969; Dickerson & Willham, 1983; Lush, 1948; Stonaker, 1973; Willham, 1970) and its expression in economically important characters (Tess & Davis, 2002; Willham, 1972) in grazing cattle production systems (Bunning et al., 2019; Long, 1980; Neville, Utley, & McCormick 1985; Ordóñez et al., 1974; Ordóñez, 1985; Plasse, 1983), as well as the effects of inbreeding (Dickerson, 1973; Lopez-Fanjul, 1974), crossing over and genetic recombination on the retention of heterosis (Koch, Dickerson, Cundiff, & Gregory, 1985; Sanders, Key, Riley, & Lunt, 2005), resulting from experiments mostly performed in a temperate climate and its application in the tropics (Madalena, 2001). It reviews the conditions (Verde et al., 2007; Depablos et al., 2010) and management practices of proven technical and economic feasibility (Arriaga, 2010; Camaripano et al., 2011; Depablos et al., 2009; Ordóñez, 1990; Pierre & Camaripano, 2009; Tejos, 2014) required to achieve the expression of genetic potentials. Finally, the requirements, advantages, and disadvantages of two alternative crossing systems are formulated (Cartwright & Fitzhugh, 1972; Cartwright, Fitzhugh, & Long, 1975; Ordóñez, 1975; Plasse, 2000; Plasse, Bauer, Galdo, & Verde, 2005a; Plasse, Bauer, Galdo, & Verde, 2005b), to the rotational system to finally compare them and recommend accordingly. The sources consulted were extracted from the mostly American scientific literature, in printed or electronic journals, memories of scientific meetings, and theses.
Review and discussion
The criteria to compare the advantages or disadvantages of the alternative mating systems proposed should consider several aspects: (1) Expected heterosis and heterosis retention are the most important; (2) Flexibility to altered management, or market conditions or unexpected performance of the cross, resulting in the loss of resources, time, and efforts; (3) The simplicity of management; (4) Previous experiences in similar conditions; (5) Interference with selection for additive traits; (6) Minimum population size.
Composites
Composite development may be indicated when heterosis is essential, if initial unfavorable recombination effects are negligible, when there are new objectives or altered management conditions, and in areas where the simplicity of the breeding program is essential. Zebu, some exotic breeds, and native breeds have been used in some instances for developing Composites in the tropics (Canchin and Montana in Brazil, Bonsmara in South Africa, Droughtmaster in Australia, Santa Gertrudis, Bradford, Brangus, and Charbray in the Southern United States).
(1) Management: Synthetic breeds will contribute to more straightforward management requirements. Kowalsky (2020) described the Venezuelan cattle herd structure: 94,6 % are smallholdings, 87% own less than 30 crossbred cows. They are well-served with just one bull. As in any pure stock, sires of similar genetic composition are used. Thus, the number of pastures required will be minimal, particularly favorable to small ranchers.
(2) Selection for additive traits: Lopez-Fanjul (n.d.) has shown that when heritabilities have been estimated in composites, their value was not found to differ from those in their parental breeds. But, selection intensity will be limited in the first generations, if inbreeding is to be kept at minimal levels as well as a need for increased numbers as the breed develops.
(3) The exploitation of heterosis and heterosis retention: Heterosis, the difference between the average performance of a cross and the average performance of the parents, has two main components. The first and most important is the result of differences in gene frequencies between the two parental populations and the degree of dominance present. Willham (1970) has shown how heterosis at a single locus in the F1 is equal to ∆ p 2 d, where ∆ 𝑝 is the difference in the gene frequency between the two parental populations, and 𝑑 is the degree of dominance. After the first filial generation F1, the gene frequencies remain constant according to the Hardy-Weinberg equilibrium, then the amount of heterosis remains constant in the successive generations. As the gene frequencies in the F1 becomes intermediate between the parental gene frequencies, the degree of heterosis is equal to ½ ∆ 𝑝 2 𝑑; meaning that half the original heterosis is retained in successive generations. In general, if n breeds are utilized in the Composite development, 1/n of the original heterosis is lost, or (n-1)/n is retained. The second component of heterosis is due to favorable epistatic combinations of linked genes in the repulsion phase (inherited separately), which had been fixed by selection or isolation in the parental breeds. Under interse mating, successive crossing over and recombination will tend to destroy those linked groups. After two or three generations, all these epistatic effects will probably be lost. When the epistatic effects are significant, loss of heterosis will be larger than what is theoretically expected. Koch et al. (1985) result supports the hypothesis that heterosis effects of crosses among Bos taurus breeds, for traits related to growth and size as well as for reproductive and maternal traits, can be accounted for by the dominance effects of genes. Nevertheless, strengthen that large-scale comprehensive experiment is needed to estimate retention of heterosis in advanced generations of interse mated composite populations with contributions by both Bos taurus and Bos indicus breeds. Indeed, Sanders et al. (2005) present additional questions regarding the validity of the dominance model for the prediction of heterosis and heterosis retention for reproductive and maternal traits in Bos indicus x Bos taurus females. Heterosis retention estimates for the traits of interest were found to be lower than expectations of the dominance model for some groups and higher than expectations of the dominance model for other groups. Plasse et al. (2005a) suggested that line formation was difficult since crosses among tropical breeds under conventional management did not exceed the best parental breed. Madalena, (2001) concluded that in Bos taurus and Bos indicus crosses for milk, the performance of the F1s has been much higher than the other crosses, followed by the rotational crosses, while the "bimestizos" (in Brazil, daughters of mestizo parents) have had very poor performance. The relative economic performance of F1, rotational cross, new breed (Composite), and Holstein were, respectively, 100, 59, 30, and 21, in privates farms.
(4) Minimum population numbers: Lopez-Fanjul (1974) states that the primary advantage of increased heterozygosity can be "squandered" by renewed inbreeding unless large population numbers are kept. How large should the experimental herd be to keep inbreeding at a low level can be estimated with some approximation.
The rate of inbreeding Δ𝐹 is a function of effective population size. Lush (1948) showed how Δ𝐹= 1 2𝑁𝑒 where 𝑁𝑒 is the effective population size and Δ𝐹 is the variation in inbreeding per generation. When there is a different number of males and females, 𝑁𝑒 becomes 1 𝑁𝑒 = 1 4𝑁𝑚 + 1 4𝑁𝑓 , where (ec1)
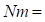 number of males and
number of males and
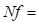 number of females (ec2). The rate of inbreeding Δ𝑓 per generation can be rewritten as Δ𝑓= 1 8𝑁𝑚 + 1 8𝑁𝑓 . As population size becomes larger, Δ𝑓 becomes smaller, and the generation with the smallest number has the most significant effect. However, as the population size increases, the previous inbreeding is not eliminated but remains where it was before the increase in number because new inbreeding is reduced. Thus, the first three or four generations, while the Composite is being tested, are the most important ones in terms of inbreeding. More than ten to twelve unrelated bulls and two hundred and fifty to three hundred cows are required per generation if less than one percent increase in inbreeding per generation is desired, assuming random mating, which is not the case. Even if selection and further assortative mating are used, the above holds because Lush, (1948) has shown them to be almost powerless in changing heterozygosis. Selection will be limited to the extent that more sires per generation will have to be used, which will reduce intensity. 𝑁𝑚=𝑝 ×𝑁, where 𝑁 is the number of males reaching reproductive age and 𝑝 the proportion saved. If selection intensity 𝑖= 𝑍 𝑝 , for i to be large, 𝑝 must be small, but if 𝑝 is small, Nm will also be small in the Composite, making Δ𝑓 large. Then the advantage of more considerable additive variance can be neutralized by the requirement of keeping inbreeding at a low level.
number of females (ec2). The rate of inbreeding Δ𝑓 per generation can be rewritten as Δ𝑓= 1 8𝑁𝑚 + 1 8𝑁𝑓 . As population size becomes larger, Δ𝑓 becomes smaller, and the generation with the smallest number has the most significant effect. However, as the population size increases, the previous inbreeding is not eliminated but remains where it was before the increase in number because new inbreeding is reduced. Thus, the first three or four generations, while the Composite is being tested, are the most important ones in terms of inbreeding. More than ten to twelve unrelated bulls and two hundred and fifty to three hundred cows are required per generation if less than one percent increase in inbreeding per generation is desired, assuming random mating, which is not the case. Even if selection and further assortative mating are used, the above holds because Lush, (1948) has shown them to be almost powerless in changing heterozygosis. Selection will be limited to the extent that more sires per generation will have to be used, which will reduce intensity. 𝑁𝑚=𝑝 ×𝑁, where 𝑁 is the number of males reaching reproductive age and 𝑝 the proportion saved. If selection intensity 𝑖= 𝑍 𝑝 , for i to be large, 𝑝 must be small, but if 𝑝 is small, Nm will also be small in the Composite, making Δ𝑓 large. Then the advantage of more considerable additive variance can be neutralized by the requirement of keeping inbreeding at a low level.
In conclusion, a large population is a condition for full utilization of heterosis resulting after crossing different breeds for the development of Composites. The cost of keeping such a large population can limit the effectiveness of this proposal.
(5) Flexibility against risk: The success or failure of the Composite is determined at the precise moment of the population closure. Sampling errors, loss of heterosis due to renewed inbreeding, or recombination, the variation in prevailing conditions (environment or market), or unexpected performance of the Composite may result in the loss of resources, time, and efforts. If, after three or four generations, the results are unsatisfactory, everything will have to start all over. The literature contains just the successful trails, while many failures stay undisclosed to the public. This risk factor must be considered before any breed development is begun. Plasse (2000) concludes, "The author is not aware of any information in the current Latin American scientific literature that would allow him to responsibly recommend this system (composite) to livestock practice." While Leachman (2000) proposed "composed populations of several original breeds, permanently open to new genetic contributions."
3.2 Modified Absorption
A second alternative to rotation system could be one that combines the simplicity of the purebred mating, exploits heterosis in successive generations with the versatility of totally open herds. Such a mating system was empirically purposed in previous papers (Ordóñez, 1975, cited by Ordóñez, 1990) based on earlier work by Cartwright & Fitzhugh (1972) and Koger (1973). A schematic representation of the mating system is depicted in figure 1.
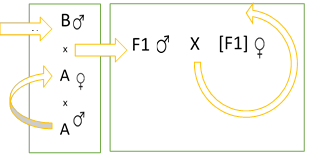
A proportion of purebred female of Breed A (adapted to the environment) is mated to sires of Breed B. The F1 progeny of that mating, bulls, and heifers, are removed from the purebred herd and incorporated into the commercial herd. F1 sires are mated to the commercial cows, originally of Breed A, in successive generations until all the females in the commercial herd are of genetic composition like the F1. The modified F1's is represented within brackets [F1]. The female progeny from the commercial herd are kept as replacements, while the males go to market. In any generation, new unrelated F1 sires are used. It can be said that the mating system tries to absorb or grade up to [F1]. The expected composition of successive generations at equilibrium has been developed with that of the rotation to allow comparisons.
Rotation: First, let us call
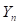 the (ec3) fraction of genes from the parental breed in the nth generation and
the (ec3) fraction of genes from the parental breed in the nth generation and
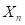 the fraction of genes from the maternal grandsire in the same generation of any progeny of rotation mating.(ec4)
the fraction of genes from the maternal grandsire in the same generation of any progeny of rotation mating.(ec4)
Then:
Or
but 𝑌 0 =1 in the purebred population, then in general:
while, (1)
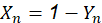
Table 1 summarizes the application of this general formula to fill the genetic composition of the herd in the successive generations of rotational crosses.
Modified [F1]: By analogy in Modified [F1] 𝑌 𝑛 is the fraction of genes from the newly introduced breed carried by the progeny in generation n; then in general (2)
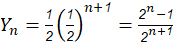
Table 2 summarizes the application of these general formulas to fill the genetic composition of the herd in the successive generations of "Modified [F1]".
As can be seen from table 2, the genetic composition of the population at equilibrium resembles that of the first generation of interse mating. The properties of such a population must be studied in more detail.
(1) The simplicity of management: At the commercial level, the system performs as simply as does any pure breeding system. Only one breed of sires is used, allowing for entire herds in the small operations. The system also provides for their replacement females. Still, F1 sires must be provided from elite herds located at accessible areas, where AI increases the ability to capitalize on genetic progress in the progenitor breeds, under the prevailing conditions.
(2) Flexibility: It is one additional advantage of the proposed system. Change in the exotic breed due to adjustment of the performance of the products following changes in environmental conditions, the market, and the goals is relatively simple and in no way affects the development of the system, reducing the risk mentioned above. The flexibility available through crossbreeding to match performance levels to specific conditions must also be considered a real advantage (Long, 1980).
(3) The adaptability of the parents: As has been indicated, a primary consideration for crossbreeding is the possibility of combining useful traits quickly in an individual. The main objective of using crossbreed bulls is to combine in an animal the adaptability of the tropical breed and the improved performance of the exotic, hoping to obtain an animal able to work in adverse conditions, but that simultaneously transmits to his progeny desired genes for beef production. F1 Zebu x European bulls have proven to be suitable as mentioned by Neville et al. (1985) on "the comparison of SB (purebred) and F1 bulls for reproductive and progeny performance… there were no differences (P>.05) among the four sire groups for the proportion of cows exposed that had a calf, had a live calf or weaned a calf." The adaptability of the female, on the other hand, is guaranteed if their genetic potential for mature size and particularly for milk production, is maintained within limits coupled with the level of nutrition (Plasse, 2000). As stated by Willham (1972), "If low-quality roughage that can be harvested only by the cow is considered, milk production in excess of growth demands by the calf seems of little economic value." The cow adaptability depends, to a large extent, on the exotic breed, which is chosen to be incorporated in the population.
Table 1 : Genetic Composition in Successive Generations of Rotational Crosses.
| Generation | 𝑌 𝑛 𝑋 𝑛Genetic composition of Females of Herd A* | Genetic composition of Females of Herd B* | ||
| 0 | 1 | 0 | 1B:0A | 1A:0B |
| 1 | 1 2 1 21B:1A | 1A:1B | ||
| 2 | 3 4 1 43B:1A | 3A:1B | ||
| … | … | … | … | … |
| n | 1+ 𝑖=1 𝑛 −2 𝑖 −2 𝑛 1− 𝑌 𝑛 𝑌 𝑛 𝐵: 𝑋 𝑛 𝐴 𝑌 𝑛 𝐴: 𝑋 𝑛 𝐵||||
| … | … | … | … | … |
| ꝏ | 2 3 1 3 2 3 𝐵: 1 3 𝐴 2 3 𝐴: 1 3 𝐵||||
| * A and B are breeds of sires mated to those females. |
Table 2: Genetic Composition in Successive Generations of "Modified [F1]".
| Generation |
|
|
Genetic composition of females | Genetic composition of sires |
| 0 | 0 | 1 | 1A:0B |
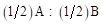
|
| 1 |
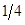
|
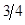
|
3A:1B | |
| 2 |
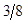
|
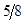
|
3B:1A |
|
| … | … | … | … | … |
| n |
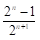
|
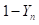
|
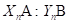
|
|
| … | … | … | … | … |
| ꝏ |
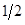
|
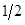
|
|
Table 3 : Some comparisons between the main systems.
| Mating System | % Crossbreed | % Heterosis of individual | Sire adaptability | Sire value | Management simplicity | |||||
| Calf | Cow | Calf | Cow | Sire | ||||||
| Purebred | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | ||
| Rotation | 100 | 100 | 67 | 67 | 0 | <1 | >1 | <1 | ||
| F1 x [F1] | 88 | 83 | 50 | 50 | 100 | 1 | 1 | 1 | ||
(4) The efficiency - the ability to produce an output without waste: As indicated by Cartwright (1970), rather than individual performance, the whole system performance must be considered when comparisons between systems must be made. Following the scheme indicated by Cartwright et al. (1975), some comparisons between the main alternatives made in a previous paper, are included here because of their importance. Although the rotational cross seems to be more useful for exploiting hybrid vigor, the last four columns of table 3 can make a difference in the profitability of the proposed system.
(5) The exploitation of Heterosis: Following Willham's (1970) approach to genetic consequences of crossbreeding, some properties of such a population can be estimated. Considering a single locus with two alleles L1 and L2 and two breeds A and B, that have gene frequencies p:q and p+Δp:q-Δp.
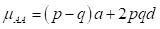
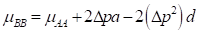
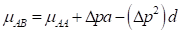 ; where µ
AB
is mid parent value,
; where µ
AB
is mid parent value,
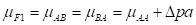
where µ F1 is the mean of the reciprocal crosses µ A.B and µ B.A; a is the average effect of a gene substitution, and d is the dominance deviation from the homozygote average.
Then, heterosis is defined as the difference between the mean of the cross and the average of the parents and is equal to:
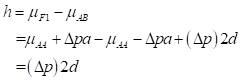 (3)
(3)For this locus to show heterosis, it is required both a difference in allele frequencies between the populations (Δp ≠ 0) and positive dominance (d > 0).
Then, the genetic array of the crossbreed F1 sire is
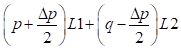
and the mean of the first-generation crossing A females and F1 males can be developed
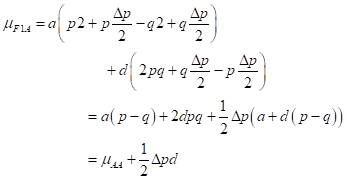 then, heterosis:
then, heterosis:
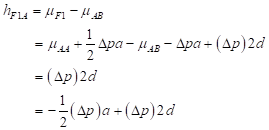
We have estimated
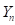 as the fraction of genes from the newly introduced breed in the progeny of generation n. This same fraction remains of the difference between gene frequencies at the locus level. Then in general, means and heterosis from the mating of F1 sires with the crossbred females of generation n can be calculated as follows:
as the fraction of genes from the newly introduced breed in the progeny of generation n. This same fraction remains of the difference between gene frequencies at the locus level. Then in general, means and heterosis from the mating of F1 sires with the crossbred females of generation n can be calculated as follows:
Genetic array:
Males:
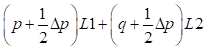
Females:
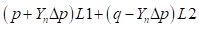
Genotypic array
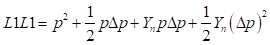
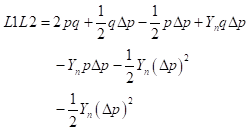
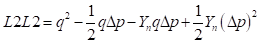
Heterosis:
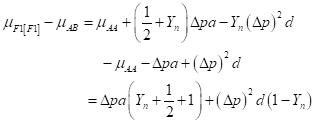
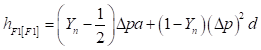 (4)
(4)
The level of heterosis depends on the fraction of the squared difference in gene frequency remaining and the degree of dominance minus the additive component of the differences.
By analogy, table 4 summarizes the application of these general formulas to fill the gene frequencies and expected heterosis of the herd in the successive generations of "Modified [F1]" cross.
The expected heterosis from the system in equilibrium is equal to that of an interse mating. However, it is necessary to indicate that while synthetics reach their maximum level of heterosis in the first generation of crosses, Modified Absorption to [F1] behaves in the opposite direction, the level of heterosis increases in successive generations as the coefficient of Δp increases. That can be a disadvantage if maximum production potential is desired rapidly, but it could be an advantage if the selection is applied simultaneously. It also gives time to the breeder to provide the appropriate environment required to match the new genotype tested. On the other hand, when epistatic effects are essential components of heterosis, F1 sires can make partial use of this advantage. If recombination is low enough and selection is applied, the initial disequilibrium between repulsion and the coupling phases may persist for several generations with the additional heterotic advantage. Furthermore, as heterosis in sire reproductive performance is important, it could represent an additional advantage of this system over the rotation.
(6) Loss of heterosis due to renewed inbreeding: As the population is maintained open, unrelated sires can be used to generate F1 sires, in every generation. Then, the probability of heterosis losses due to renewed inbreeding is small if nonexistent.
(7) Selection for the additive traits: As said before, F1 sires must be provided from elite herds where AI is available, capitalizing on the genetic progress in the progenitor breeds. Then, selection among the purebred herd that generates the F1 sires will have similar results that they would have under purebred mating. Willham (1970), discussed how selection response in crossbreds is lower than that obtained when selecting among purebreds or advanced generations of interse mating. However, the heritability of a trait is a ratio between the additive and total phenotypic variances. Although the dominance and epistatic components of variance are increased in the crossbred, the additive component is also increased. Stonaker (1973) has found that heritabilities are somewhat higher in crossbreds rather than in purebreds.
Table 4 : Expected heterosis of the herd in successive generations of "Modified [F1]".
| Generation | Gene frequency of females | Gene frequency of sires | Expected heterosis |
| 0 |
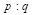
|
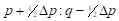
|
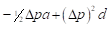
|
| 1 |
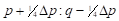
|
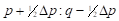
|
|
| 2 |
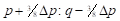
|
|
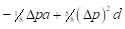
|
| … | … | … | … |
| n |
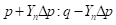
|
|
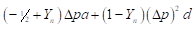
|
| … | … | … | … |
| ꝏ |
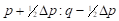
|
|
Since crosses are generally less variable, this higher heritability is probably the result of a smaller environmental and a more significant genetic contribution to the difference. Additionally, Sires in Breed B might be selected mainly on additive merit based on their progeny from cows of Breed A.
If nonadditive effects contributed to their ranking on the EBV, then the Sires selected in Breed B would be chosen in part for their genetic differences from Breed A. Thus, the long-term effects of selection in the two breeds would be for genetic divergences, and hence the enhancement of heterosis between them.
4 Comparison of alternative mating systems
4.1 Similarities
Comparing Composites and Modified [F1], it becomes evident that (1) both mating systems generate female replacements throughout the herd and (2) effectively exploit heterosis to a similar extent, given they are made of the same number of breeds. (3) 100% of the cowherd as well as of the calves marketed are crossbreds so, (4) if meticulously designed, females and males could be fully adapted to the conditions under which they will strive. (5) Minimum number of breeding pastures required is one.
4.2 Differences
However, some differences need to be highlighted: Modified [F1] overcome three fundamental requirements: (1) The use of F1 sires impede loss of heterosis due to renewed inbreeding, expand heterosis retention, do not interfere with selection for additive traits, and capitalize genetic progress in the parental breeds. (2) There is not a minimum herd size, a valuable attribute for the small rancher. (3) The flexibility available, through a change in the exotic breed, to match genetic potential to specific conditions of the environment, market, and goals.
Finally, a valid criterion to compare the advantages or disadvantages of the alternative matting systems is "previous experiences” in similar conditions. There are no experiences with the Modified [F1], but, as mentioned before, the literature contains just the successful trails while many failures stay undisclosed to the public. Plasse (2000) was very brave to admit, after several attempts, that he was not aware of any information that would allow him to recommend composites to livestock practice. So, in the absence of previous experience with the Modified [F1] system, more research on mating systems in beef cattle production is required.
5 Conclusions
From this review, it can be inferred that significant progress could be made in terms of adopting improved management procedures currently being used in experimental stations and elite herds. However, for the next several years, the beef cow and their lactating progeny will be relegated to marginal areas where environmental limitations or lack of adaptability prevent the expression of genetic potentials.
The main objective of using crossbreed bulls is to combine in an animal the adaptability of the tropical breed and the improved performance, hoping to obtain an animal able to work in adverse conditions, but that simultaneously transmits to its progeny desired genes for beef production and exploit heterosis.
Heterosis, the superiority of the crosses over the average of the parent breeds, improve economic efficiency in systems that exploit both individual and maternal heterosis. However, heterosis is not a parameter determined by the genotypes that intervene in the crossing; its effects are conditioned by environmental limitations or lack of adaptability that prevent the manifestation of the genetic potentials.
In general, if n breeds are utilized in the development of the Composite, 1/n of the original heterosis is lost under interse mating in successive generations. Additionally, crossing over and recombination will tend to destroy linked groups, and after two or three generations, all epistatic effects will be lost. When the epistatic effects are important, loss of heterosis will be more significant than what is theoretically expected. Furthermore, a large population is a condition for full utilization of heterosis resulting after crossing different breeds for the development of composites.
For the traits of interest, in Bos indicus x Bos taurus females, heterosis retention estimates were found to be lower than expectations of the dominance model for some groups and higher than expectations of the dominance model for other groups, which confirm that more research is needed on mating systems in B indicus crosses for beef cattle production.
The use of F1 sires able to thrive on adverse conditions, on a crossbreed population of cows in successive generations, might be a feasible way to overcome some of the Composites limitations, combine adaptability with improved performance in small herds while keeping management simple, capacity for adjustment, and adaptation to new restrictions and opportunities generated by the changing environment.

