











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Capparaceae Juss. es una familia de plantas perennes constituidas por especies de hábitos arbustivos, arbóreos y en ocasiones lianas; está constituida por 35 géneros y 440 especies, que están distribuidas en las zonas tropicales y subtropicales principalmente de Asia, África, Australia y América. Los miembros de Capparaceae son mayormente conspicuos y hasta abundantes en hábitats áridos [1, 2].
La familia Capparaceae es conocida por la diversidad de hábitos, frutos, y características florales, mismas que poseen una morfología variada, presentando tanto flores con simetría actinomórfica como zigomórfica Los caracteres florales resaltan en las Capparaceae debido a que poseen ginóforos, andróforos o androginóforos, las cuales son estructuras prolongadas que sostienen y proyectan al ovario hacia la parte más externa de la flor [1]. Estas estructuras son adaptaciones que los miembros de esta familia poseen y juegan un papel importante en la polinización.
Taxonómicamente, Capparaceae estaba dividida en dos principales subfamilias, Cleomoideae y Capparoideae, sin embargo, existen algunos autores que las consideraban grupos a nivel de familia [1]. Posteriormente, análisis filogenéticos y moleculares demostraron que Cleomoideae era parafilético y que además estaba más cercanamente emparentado con Brassicaceae que con Capparaceae, lo que eventualmente hizo que diversos autores reconocieran formalmente a Capparaceae, Cleomaceae y Brassicaceae como 3 familias diferentes [3, 4].Las relaciones filogenéticas se dilucidaron parcialmente en Capparaceae, al establecer que Capparis (género tipo de esta familia) era un clado difilético, es decir, que está compuesto por dos grandes linajes, por un lado, el clado Capparis sensu stricto, constituido por las especies del “Viejo Mundo” y un segundo clado que agrupaba a las especies del “Nuevo Mundo” (América)[5].
En el Neotrópico su distribución frecuentemente es notoria en hábitos xerofíticos, es decir en bosques secos hasta desiertos. Sin embargo, Capparaceae también puede ser encontrada en bosques húmedos. En la actualidad las Capparaceae del Neotrópico poseen 22 géneros y 110 especies [6, 7, 8, 9]. La presencia y predominancia de esta familia en bosques secos sugiere una importancia evolutiva y en conservación de estos hábitats en el Neotrópico, debido a que las condiciones ambientales y climáticas de esta región han contribuido a generar considerables endemismos.
Después de la reclasificación de los géneros que pertenecían al clado Neotropical reconocido como Capparis sensu lato [5] una revisión de la morfología de las estructuras florales y sus caracteres es necesaria. En el presente trabajo se plantea realizar un análisis comparativo de la estructura y de los caracteres florales de los géneros de Capparaceae de Ecuador, determinando los caracteres florales y estableciendo las similitudes y diferencias existente entre cada género.
Materiales y Metodos
Se realizó un análisis comparativo de los 11 géneros registrados en el Ecuador entre enero y mayo del 2022. Los caracteres florales fueron recopilados de una revisión de literatura científica disponibles en: Journal of the Botanical Research Institute of Texas, Brittonia, Harvard Papers in Botany, Phytotaxa, Systematic Botany y Revista Científica de Ciencias Naturales y Ambientales entre los cuales están artículos del tipo descripciones de géneros, revisiones y restablecimiento de taxones de la familia Capparaceae en el Ecuador. Además, se realizaron observaciones de docenas de imágenes de alta resolución de flores en campo. Esto se complementó con revisiones de los especímenes conservados en el Herbario GUAY.
El análisis fenético fue llevado a cabo en base a una matriz básica de datos (MBD) previamente desarrollado en base al análisis comparativo inicial de los caracteres florales. Se designaron las 11 unidades taxonómicas operativas correspondiente a los géneros presentes de la familia Capparaceae en Ecuador; posteriormente se determinaron los caracteres válidos con los estados de cada carácter mediante claves taxonómicas y descripciones de géneros [10, 11, 12]. Todos los caracteres florales y ecológicos de cada género se resumen en la matriz básica de datos, presentados en la Tabla 1 (A) y Tabla 1 (B)
Tabla 1: (A) Matriz básica de datos de los caracteres florales y ecológicos utilizada para el análisis fenético.

Nota: La continuación de la Tabla 1 (A) se detalla en la Tabla 1 (B).
Tabla 1: (B) Matriz básica de datos de los caracteres florales y ecológicos utilizada para el análisis fenético.

Nota: A: Colicodendron B: Beautempsia C: Capparidastrum D: Morisonia E: Cynophalla F: Preslianthus G: Capparicordis H: Neocalyptrocalyx I: Crateva J: Neocapparis K: Steriphoma
Los coeficientes de similitud fueron estimados mediante la herramienta Simqual incluida en el software NTSYS-pc, el análisis de agrupamiento fenético se ejecutó con el “Agrupamiento jerárquico y anidado aglomerativo secuencial” (Sequential Agglomerative Hierarchical and Nested clustering por sus siglas en ingles SAHN) herramienta que también está incluido en el software NTSYS-pc.
Resultados y discusión
Determinación de las características florales de los 11 géneros de la familia Capparaceae de Ecuador
A continuación, se describen los resultados de las respectivas descripciones de los 11 géneros de la familia Capparaceae presentes en Ecuador.
Colicodendron. Arbustos o árboles. Inflorescencias terminales, subterminales y/o axilares, principalmente panículas o racimos en espigas. Pubescencia estrellada. Brácteas lineales. Brote floral por lo general globoso-elipsoide, cáliz valvado, estivación cerrada hasta antes de la antesis. Flores zigomórficas. Cáliz de cuatro sépalos uniseriados, gamosépalos. Sépalos monomórficos, triangulares a lanceolados. Corola de cuatro pétalos, dialipétala, estivación cerrada (imbricada o torsiva). Pétalos con tonalidades amarillas blanquinosas, obovados, nervios longitudinales. Hipantio presente, recubierto por un plato nectario con cuatro glándulas o una copa nectaria trilobulada; glándulas nectarias dentado-lobuladas. Androginóforo corto y descentrado. Estambres 14 a 60. Ginóforo presente. Ovario ovoide a elipsoide, estigma sésil y truncado. Habita comúnmente en bosques secos de la costa. Florece durante la noche.
Beautempsia. Arbustos o árboles. Inflorescencias subterminales y opuestas a las hojas distales, dispuestas en racimos. Pubescencia estrellada. Brácteas lineales. Prefloración comúnmente globosaelipsoide, cáliz valvado, estivación abierta. Flores zigomórficas. Cáliz con cuatro sépalos uniseriados, gamosépalos. Sépalos monomórficos, tetralobulados a ligeramente tetracrenados y subtruncados. Corola con 4 pétalos, dialipétalo, estivación cerrada. Pétalos color crema, obovados, patrón de los nervios longitudinales. Hipantio presente, recubierto por un plato nectario con cuatro glándulas mayormente libres y fusionadas a la base; las glándulas son deltoides. Androginóforo corto y grueso coronado por un pseudotorus digitado. Estambres 16 a 18. Ginóforo presente. Ovario ovoide a elipsoide, estigma sésil y capitado. Habita en bosques secos de la costa. Florece durante el día.
Capparidastrum. Arbusto o árbol. Inflorescencia terminal dispuesta en racimos. Indumenta simple o incluso glabrescente. Brácteas deltoides. Brote floral de globoso, obovado a elipsoide, valvado e imbricado con estivación abierta. Flores actinomórfica. Cáliz con cuatro sépalos, uniseriados, dialisépalos. Sépalos monomórficos, deltoides a hemiorbiculares. Corola de cuatro pétalos, dialipétala, estivación cerrada. Pétalos con tonos blancos a cremosos, ovados a ovado-elípticos, nervios longitudinales. Receptáculo plano y disciforme. Cuatro glándulas subglobosas dorsoventralmente comprimidas, episépalas, con tonalidades variadas que van desde el rojo púrpura, rosado a verde claro. Androginóforo ausente. Estambres 22 a 60. Ginóforo presente. Ovario de ovoide, sublanceolado, elipsoide a subcilíndrico, estigma sésil y truncado o hemisférico. Habita predominantemente en bosques húmedos en ambos lados de los Andes, algunas especies habitan en bosques secos de la costa. Florece durante el día y la noche.
Morisonia. Árbol. Inflorescencia principalmente ramiflora o subterminal dispuesta en racimos. Tricomas estrellados a estrellado-equinado. Brácteas lineales. Brote floral globoso ovado, cáliz valvado, reticulado y estivación cerrada hasta antes de la antesis; el cáliz se rompe en segmentos sepaloides irregulares al entrar a la antesis. Flores actinomórficas. Cáliz con dos a cuatro sépalos uniseriados no definidos, gamosépalos. Sépalos monomórficos, comúnmente triangulares. Corola con 4 pétalos, dialipétala y estivación cerrada. Pétalos verde amarillentos, ovados, oblongos a obovados con nervios longitudinales y nervadura reticulada en los márgenes. Hipantio presente, recubierto por una copa nectaria con cuatro glándulas dentadas. Androginóforo corto, ensanchado en el ápice. Estambres 17 a 24. Ginóforo presente. Ovario ovoide a globoso y costulado, estigma sésil y cupuliforme. Habita en bosques secos de la costa. Florece durante el día.
Cynophalla. Arbustos o árboles. Inflorescencias terminales dispuestas en panículas, racimos o corimbos. Tricomas simples o glabro. Brácteas deltoides. Prefloración globosa-elipsoide, cáliz con sépalos decusados, estivación abierta. Flores zigomórficas. Cáliz con 4 sépalos distintivamente biseriados, dialisépalos. Sépalos dimórficos, el par externo con mayores dimensiones que los internos, ambos pares de sépalos hemiorbiculares. Corola cuatro pétalos, dialipétala, estivación cerrada. Pétalos blancos a blancos verdosos, obovados, con nervadura longitudinal. Receptáculo plano en forma de disco con cuatro glándulas nectarias planas o escuamiformes insertas. Glándulas nectarias extraflorales en la inflorescencia de 1 a 3, cupuliformes a ciatiformes. Androginóforo corto y ancho. Estambres 28 a 50. Ginóforo presente. Ovario subcilíndrico, estigma sésil y capitado. Habita en bosques secos y húmedos de la costa. Florece durante la noche.
Preslianthus. Arbustos o árboles. Inflorescencias terminales dispuestas en racimos o corimbos. Pubescencia estrellada. Brácteas no observadas. Brote floral globoso, cáliz valvado con estivación abierta. Flores zigomórficas. Cáliz de cuatro sépalos uniseriados hasta sépalos apenas biseriados, dialisépalos. Sépalos monomórficos, ovados. Corola de cuatro pétalos, dialipétala, estivación abierta. Pétalos verdes a verde amarillentos, ovados a amplioelípticos, nervios longitudinales. Receptáculo plano y disciforme con cuatro glándulas escuamiformes insertas. Androginóforo corto presente. Estambres 14 a 40. Ginóforo presente. Ovario ovoide, estigma sésil y capitado. Habita en bosques húmedos de la Amazonia. Florece durante el día.
Capparicordis. Arbustos. Inflorescencias subterminales y opuestas a las hojas distales en forma de racimos, corimbos o incluso en flor solitaria. Pubescencia estrellada. Brácteas lineales. Prefloración comúnmente elíptico-elongada, cáliz valvado, estivación cerrada hasta antes de la antesis. Flores zigomórficas. Cáliz de cuatro sépalos uniseriados, dialisépalos. Sépalos monomórficos, lanceolados. Corola de cuatro pétalos, dialipétala, estivación cerrada. Pétalos con tonalidades amarillentas, oblanceolados, nervios pinnatinervios. Hipantio presente, revestido por una copa nectaria con 4 glándulas en forma de apéndices filiformes o deltoides. Androginóforo presente. Estambres 6 a 8. Ginóforo presente. Ovario globoso, elíptico y elongado a ovoide, estigma sésil y casi truncado. Habita en bosques secos de la costa. Florece durante el día y en la noche.
Neocalyptrocalyx. Árboles. Inflorescencias axilares, supráxilares hasta subterminales, principalmente corimbos, racimos o flor solitaria. Pubescencia estrellada. Brácteas lineales. Brote floral globoso, globoso-ovado a globoso-elipsoide, cáliz con sépalos decusados (par exterior valvado-connado), estivación cerrada con el par de sépalos internos no visibles hasta antes de la antesis. Flores actinomórficas. Cáliz 4 sépalos distintivo biseriado, dialisépalos. Sépalos dimórficos (par externo de mayores dimensiones que los internos); par externo amplio-ovado y par interno lanceolado-oblongoide. Corola de cuatro pétalos, dialipétala, estivación cerrada. Pétalos blancos, obovados con nervadura longitudinal. Receptáculo plano en forma de disco con cuatro glándulas nectarias escuamiformes insertas. Androginóforo presente. Estambres 7 a 37. Ginóforo presente. Ovario ovoide-elíptico y longitudinalmente costado, estigma sésil y casi truncado a capitado. Habita en bosques húmedos de la Amazonía. Florece durante la noche.
Crateva. Arbustos o árboles. Inflorescencias terminales dispuesta en racimos corimbiformes. Glabrescente. Brácteas no observadas. Brote floral ovado, cáliz valvado, estivación abierta. Flores zigomórficas. Cáliz con cuatro sépalos uniseriados, dialisépalos. Sépalos monomórficos, lanceolados. Corola de cuatro pétalos, dialipétala, estivación abierta. Pétalos blancos o incluso púrpura, unguiculados, nervios longitudinales. Hipantio presente, con un plato o copa nectaria y borde engrosado. Androginóforo corto y grueso. Estambres 8 a 50. Ginóforo presente. Ovario globoso, ovoide, elíptico a linear, estigma sésil a subsésil y capitado. Habita en bosques húmedos de la costa y Amazonía. Florece comúnmente durante el día.
Neocapparis. Árboles. Inflorescencias terminales dispuestas principalmente en racimos. Inflorescencia con tricomas simples o glabro. Brácteas lineales. Prefloración globoso-ovado, cáliz valvado, estivación abierta. Flores actinomórfica. Cáliz de cuatro sépalos uniseriados, dialisépalos. Sépalos monomórficos, ovados a deltoides. Corola de cuatro pétalos, dialipétala, estivación cerrada. Pétalos blancos, obovados, nervios longitudinales. Hipantio presente con 4 glándulas nectarias escuamiformes insertas. Androginóforo presente. Estambres 30 a 63. Ginóforo presente. Ovario esférico, ovoide hasta ampliamente elíptico, estigma sésil y capitado. Crece en bosques secos de la costa. Florece durante la noche.
Steriphoma. Lianas leñosas. Inflorescencias terminales dispuestas en racimos. Pubescencia estrellada. Brácteas lineales. Brote floral oblongo-elipsoide hasta clavado, cáliz valvado, estivación cerrada hasta antes de la antesis. Flores actinomórficas. Cáliz de 2 a 3 lóbulos uniseriados, gamosépalos. Sépalos monomórficos, triangulares a lanceolados. Corola de cuatro pétalos, dialipétala, estivación cerrada. Pétalos con tonalidad blanco-cremosa, oblanceolados, nervios longitudinales. Hipantio presente, recubierta por un plato nectario y tejido glandular en forma de anillo. Androginóforo presente. Estambres 6. Ginóforo presente. Ovario ampliamente elíptico a cilíndrico,estigma sésil y capitado. Habita en bosques secos y húmedos de la costa. Florece durante el día y la noche.
Análisis fenético (análisis de similitud y agrupamiento)
En el análisis de similitud se registró el máximo valor (0,8667) entre los géneros Colicodendron y Beautempsia, esta fuerte relación que tienen estos dos géneros simpátricos es contrastante con la diferencia entre sí que presentan en cuanto a caracteres como los tipos de inflorescencias, tamaños florales y polinizadores (insectos versus murciélagos) [12, 13]. Sin embargo, cabe destacar que ambos géneros pueden ser encontrados dentro de bosques xerofíticos de la costa hasta los semidesiertos peruanos, lo que le da un mayor sustento a la relación fenética y ecológica de los géneros. Así mismo, se estimó una maxima similitud (0,8667) entre Capparidastrum y Neocapparis; su relación cercana se explica debido a que este último género pertenecía a Capparidastrum como subgénero Pachycarpum, pero fue elevado a estatus genérico al evidenciarse una difilia de Capparidastrum subg. Capparidastrum a nivel molecular [6, 14], ambas taxas poseen relaciones morfológicas en común como el tipo de tricomas simples o glabros dependiendo de la especie, la ubicación terminal de los racimos, prefloraciones globosas, estivación del cáliz abierta, características las cuales permiten considerarlos como géneros cercanos [6, 15].
Por otra parte, Crateva y Neocalyptrocalyx mostraron la menor similitud (0,3667) entre dos géneros de la familia Capparaceae. Seguido de Steriphoma y Neocalyptrocalyx, que también mostraron una mínima similitud (0,4667). Los coeficientes de similitud de todos los pares de géneros posibles se encuentran registrados en la Tabla 2.
Tabla 2:Coeficiente de similitud entre los géneros de la familia Capparaceae.
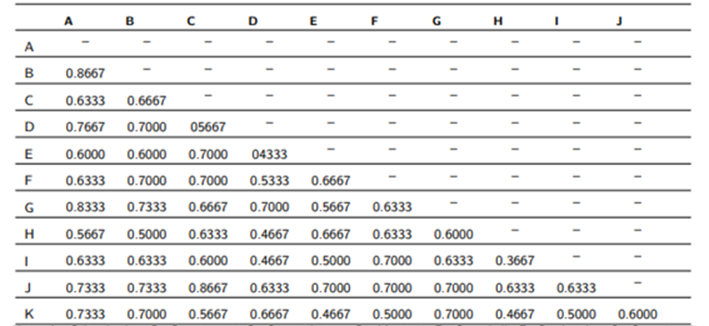
Nota: A: Colicodendron B: Beautempsia C: Capparidastrum D: Morisonia E: Cynophalla F: Preslianthus G: Capparicordis H:Neocalyptrocalyx I: Crateva J: Neocapparis K: Steriphoma
Se generaron dos cladogramas, el primero utilizó datos de caracteres florales y ecosistémicos, mientras que el segundo fue generado con datos florales, ecosistémicos y un carácter vegetal (hojas simples vs. Trifolioladas). El primer cladograma demuestra la existencia de dos clados (Figura 1). El clado ”A” formado por Colicodendron, Beautempsia, Capparicordis, Morisonia y Steriphoma mismo que comprende géneros de árboles y arbustos que presentan tricomas estrellados hasta equinados, están restringidos a los bosques de la costa de Ecuador, predominantemente desde bosques secos hasta montes espinosos tropicales, la única excepción es Steriphoma, el género más externo de este clado que posee el hábito de ser liana en Ecuador y que además se encuentra en bosques húmedos [16]. Dentro del clado A se conforma un subclado “C” conformado por géneros simpátricos.
El clado “B” está conformado por Capparidastrum junto a Neocapparis, Cynophalla, Neocalyptrocalyx, Preslianthus y Crateva. Este clado presenta dos subclados, el subclado “D” y el “E”. El primero de los subclados compuesto por los géneros Capparidastrum, Neocapparis y Cynophalla propios de hábitats costeros en Ecuador, que se encuentran ampliamente distribuidos en América Tropical, los cuales se caracterizan por ser glabros o presentar tricomas simples, cálices con estivación abierta y sépalos valvados o apenas imbricados hasta decusados, mientras que Neocalyptrocalyx, el género más externo de este subclado, es estrictamente amazónico y de las Guianas, difiere por poseer tricomas estrellados y una única estructura de cálices de estivación cerrada con los 2 sépalos externos connados que encierran a toda la prefloración hasta la antesis [11, 17]. Por otro lado, el subclado “E” está constituido por Preslianthus y Crateva los cuales componen un grupo aparte; ambos se caracterizan y son únicos en Ecuador y América Tropical por presentar prefloraciones abiertas desde muy jóvenes [18].

Nota: C: subclado “C”; D: subclado “D”; E: subclado “E”
Fig. 1: Cladograma generado mediante el análisis de agrupamiento (SHAN) tomando en cuenta caracteres florales y ecosistémicos.
El segundo cladograma comprende un grupo interno y un grupo externo (Figura 2). El grupo interno conformado por todos los géneros neotropicales caracterizados sin excepción por presentar hojas simples, este grupo denominado “clado principal” en este trabajo comprende dos subgrupos, el clado “X” conformado por Colicodendron, Beautempsia, Capparicordis, Morisonia y Steriphoma, mientras que el clado “Y” está constituido por Capparidastrum, Neocapparis, Preslianthus, Cynophalla y Neocalyptrocalyx. Por otro lado, el grupo externo está conformado por Crateva, un género Pantropical el cual es el único miembro de la familia Capparaceae en Ecuador en poseer hojas trifolioladas. Es importante mencionar que Crateva no figura dentro del clado de las Capparoides del Nuevo Mundo en los análisis moleculares [14, 19]. A pesar de esto, el género está presente dentro de América y su distribución Pantropical indica una conexión con las poblaciones y especies de este género en el Viejo Mundo. La situación de Crateva se ve reflejada en este trabajo dentro del segundo dendograma, debido a que, la inclusión de una característica vegetativa (el tipo de hoja) en el análisis fenético permitió diferenciarla de los demás géneros neotropicales, estando Crateva completamente separada del clado principal de las Capparaceae Neotropicales.
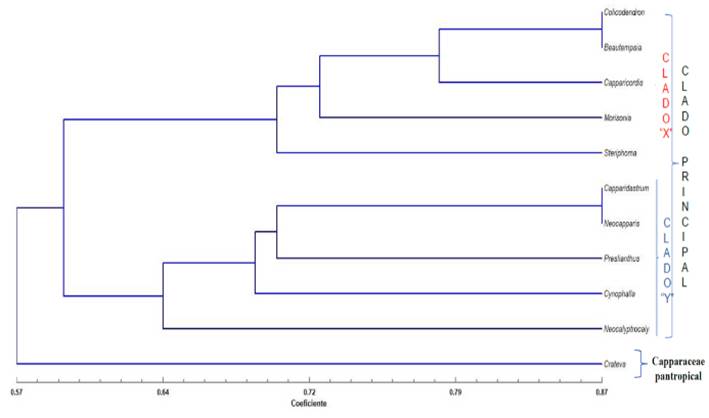
Conclusiones
El estudio comparativo de los caracteres florales según los resultados obtenidos en ambos cladogramas muestran la formación de un clado conformado por Colicodendron, Beautempsia, Capparicordis, Morisonia y Steriphoma los cuales en su mayoría comparten caracteres florales y demuestran que los caracteres con mayor relevancia en el emparejamiento y formación de clados fueron: el tipo de pubescencia, estivación del cáliz, estivación de la corola y la presencia de hipantio. Sin embargo, cabe destacar que la adición de caracteres vegetativos como el tipo de hojas y de los patrones ecosistémicos dieron lugar a la generación de dos árboles fenéticos que tienen similitudes con estudios realizados con análisis moleculares.

