











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La familia Haemulidae está representada por especies nectobentónicas que se distribuyen por los océanos Pacífico, Índico y Atlántico (From, 1986). Uno de sus principales representantes es Haemulopsis axillaris, distribuida en las costas del océano Pacífico, desde México hasta el Perú (Méndez, 2001). En Ecuador estos organismos son muy frecuentados, por lo cual representan una alternativa sustentable para generar recursos económicos en base a actividades pesqueras, gastronómicas, comerciales e industriales (López, 2012).
Las principales investigaciones que se tienen de Haemulopsis axillaris, radican exclusivamente en información descriptiva de reconocimiento de la especie. Ecuador a pesar de ser uno de los principales países acogedores de estos organismos, no registra estudios científicos que permitan conocer a mayor profundidad aspectos característicos, tales como la longevidad, maduración sexual, talla, edad y fisiología, (Arancibia, 1978).
La identificación de la edad de peces en base a la interpretación de las líneas concéntricas de otolitos sagitta, es una técnica con un alto grado de efectividad (Hubbs, 1998). En Ecuador son pocos los estudios realizados en alguna especie sobre estas estructuras óseas; en Merluccius gayi en el año 2014 se estudiaron estas marcas de crecimiento impresas en los otolitos producto a los cambios de ambiente en que habita el pez o a su vez de la propia fisiología del organismo, registrándose datos y técnicas que merecen ser llevadas a revaloración de efectividad en otro tipo de especie (Tello, 2014).
La metodología de Danevig (1956), llamada también corte ventral de las cápsulas óticas (formado por tres capas denominadas: periostal, endostal y encondral), es uno de los procedimientos con mayor efectividad para la extracción de otolitos de tipo sagitta (saculus) (Bravo, 2016). En el 2016, éste protocolo fue aplicado en el estudio de otolitos de peces de la familia Carangidae del Pacífico ecuatoriano (Bravo, 2016).
El uso de peróxido de hidrógeno para el blanqueamiento de otolitos de Merluccius gayi, es una técnica efectiva para su clarificación y posterior lectura, además demanda un bajo costo económico con respecto a otros productos químicos como el xilol que es un componente nocivo para la salud (Tello, 2014). Por lo tanto, corroborar dicho grado de efectividad en otro tipo de especie como Haemulopsis axillaris, permitirá que está técnica sea propiciada para utilizar o tomar en cuenta en todo protocolo de análisis de otolitos. La importancia pesquera y ecológica de Haemulopsis axillaris, conlleva a que la presente investigación tenga como objetivo la identificación de la edad y maduración gonadal de esta especie en base a la interpretación de las líneas concéntricas de otolitos sagitta y biometría externa e interna del organismo, iniciando de esta forma precedentes investigativos sobre estos organismos nectobentónicos, generando información relevante que puede ser usada en un futuro para establecer periodos de captura de este individuo u otros aspectos relacionados directamente al sector pesquero dada la gran aceptación comercial que tiene esta especie en nuestras costas.
Materiales y Métodos
Sitio de Estudio
Los peces Haemulopsis axillaris, fueron obtenidos en el sector pesquero “La Caleta” del cantón La Libertad (2°13'11''S; 80°54'52"W), provincia de Santa Elena, Ecuador (Fig. 1), durante el período octubre de 2017 a enero de 2018.
Ejemplares y análisis
Un total de 232 organismos de Haemulopsis axillaris (Fig. 2) fueron adquiridos para el estudio biométrico y extracción de otolitos sagitta.

La investigación comprendió aproximadamente 15 semanas, ejecutándose un total de 30 monitoreos (dos por semana), obteniendo semanalmente 16 organismos, receptados de las embarcaciones que arribaban al sector pesquero La Caleta; los peces obtenidos fueron clasificados taxonómicamente por especie, utilizando el manual para la identificación de peces óseos marino-costero del Pacífico (Steindachner, 1869) y manual de claves para identificar los peces marinos del Perú (Chirichigno, 1994). Paralelamente, se registraron los aspectos morfométricos: longitud total, ancho total; en unidades de centímetro implementando como instrumento de medición un ictiómetro; para finalmente ser colocados a refrigeración, durante lo que se denominó fase de campo.
En la fase de laboratorio, las muestras fueron transportadas semanalmente en recipientes prácticos a las instalaciones del Laboratorio de Prácticas Experimentales de la Universidad Estatal Península de Santa Elena, donde se procedió a determinar datos biológicos como masa, sexo, maduración gonadal y edad. La masa de cada organismo se estableció con el uso de una balanza digital marca Boeco en unidades de gramo
Para establecer el sexaje de cada individuo se realizó un corte longitudinal en la zona ventral del organismo, y posterior se procedió con la observación macroscópica externa, siendo así en general los ovarios tubulares, rosados y granulares, en tanto que los testículos son planos y blancos (FAO, 2012).
De forma continua se realizó una extracción gonadal con el fin de establecer el estadío de maduración sexual, realizando una revisión visual total a cada glándula genital, en base a las características externas otorgadas por la guía: descripción macroscópica de la escala de madurez gonadal del jurel Trachurus murphyi (Sánchez et al., 2013); además se utilizó el estereomicroscopio marca Boeco en objetivo 4X para observar los gametos y comprobar el estadio gonadal de cada individuo, a su vez cada gónada fue medida y pesada usando la metodología ya mencionada para establecer los datos generales de cada pez .
La edad de los peces Haemulopsis axillaris se estableció mediante la lectura de los anillos concéntricos presentes en los otolitos sagitta. Implementando la metodología de Danevig (1956), se realizó un corte transversal con una tijera de disección por delante de la aleta pectoral separando la cabeza del cuerpo, de forma continua se colocó boca abajo cada cabeza y se retiró de forma manual el exceso de tejidos, posteriormente se realizaron cortes ligeros a la parte posterior de la cavidad neurocraneal, permitiendo romperla y retirar cada uno de los otolitos sagitta (izquierdo y derecho) usando una pinza de disección.
Paralelamente las estructuras óseas fueron sometidas a una limpieza colocándolas en cajas de Petri con agua destilada, con la finalidad de eliminar los fluidos y tejidos blandos que recubrían a cada otolito; para luego ser sometidos a un aclaramiento, colocando las estructuras en tubos de ensayo de 10 ml con 1 mililitro de peróxido de hidrógeno al 30%, durante un período de aproximado de 1 minuto (Tello, 2014).
Finalmente, los otolitos fueron llevados a cajasde Petri (secas y limpias), conservando el ordenderecho e izquierdo, para su respectivo análisis ylectura de anillos concéntricos en el estereomicroscopio utilizando un aumento de 2X. Por otra parte, se tomó la medida en unidadesde centímetro del largo y ancho de cada par deotolitos (izquierdo y derecho), con ayuda de uncalibrador de Vernier (Muñoz, 2012).
Estimación de los parámetros de crecimiento
La correlación longitud-edad se realizó en base a la ecuación de von Bertalanffy L t -k(t - to) ], donde L t = L ∞ [L -e = longitud a la edad t en centímetros, L ∞ = longitud asintótica, k= coeficiente instantáneo de crecimiento, t 0 = edad teórica a la cual el pez alcanzaría la edad cero, e= base de logaritmos neperianos (Ignacio, 1986). Los parámetros de crecimiento se estimaron empleando el método de von Bertalanffy, aplicando así el diagrama de regresión lineal con los datos promedios de tallas y edades representativas, siendo así: 19,38 cm correspondientes a la edad 0+; 24,95 cm a la edad 1+ y 35,70 a la edad 2+ (Csirke, 1980), para posterior aplicar la línea de tendencia otorgando así los valores de la ecuación de la recta y= ax + b; donde y= variable dependiente, a= pendiente, x= variable dependiente y b= coeficiente de posición (Sparre y Benema, 1995)
El valor de k se calculó con k = b, mientras que a su vez el valor de t 0 se determinó con t 0 = -k/b (Bahamon, 2008), la longitud asintótica (L ∞ se calculó usando el Método de Taylor donde L ∞ = L Max /0,95; siendo L Max la longitud máxima registrada en la población (Taylor, 1958).
El gráfico de la curva de crecimiento von Bertalanffy de la correlación longitud-edad se estableció remplazando los valores de los parámetros de crecimiento en la ecuación correspondiente a este método, otorgando valores a t de 1 a 12 (edades teóricas), para esto se utilizó el programa Excel 2016.
Resultados
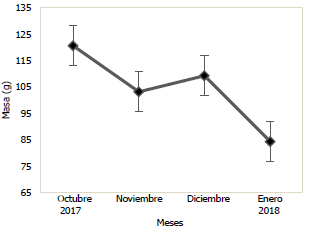
De octubre de 2017 a enero de 2018 se obtuvieron 232 organismos de Haemulopsis axillaris, 123 (53%) machos y 109 (47%) hembras (Fig. 3). Los peces presentaron una longitud promedio de 20,01 cm, con una talla mínima de 13,60 cm (enero 2018) y una máxima de 35,70 cm (octubre 2017); mientras que la masa promedio fue de 112,94 g oscilando entre 29,00 g (enero 2018) y 539,20 g (octubre 2017).
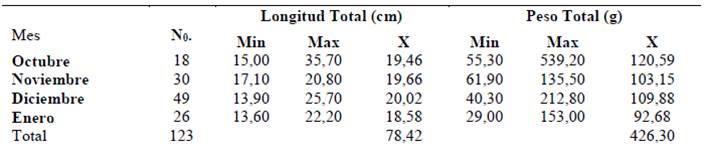
Los ejemplares machos presentaron una longitud promedio de 19,55 cm, presentando un intervalo de talla entre 13,60 cm (enero 2018) y 35,70 cm de longitud total (octubre 2017) y masa promedio total 105,36 g; de 29,00 g (enero 2018) a 539,20 g (octubre 2017) (Tabla 1).
Para las hembras el promedio de longitud total registrado fue de 20,52 cm, contemplando una talla mínima de 14,20 cm (en enero 2018) y una talla máxima de 27,30 cm (diciembre 2017), mientras que el promedio de masa total fue de 121,50 g, registrando tallas desde 56,10 g (octubre 2017) a 275,10 g (diciembre 2017) (Tabla 2)
Tabla 1 Distribución de longitud y peso de organismos hembras Haemulopsis axillaris, obtenidos en el sector La Caleta, cantón de La Libertad, entre octubre 2017 y enero 2018.
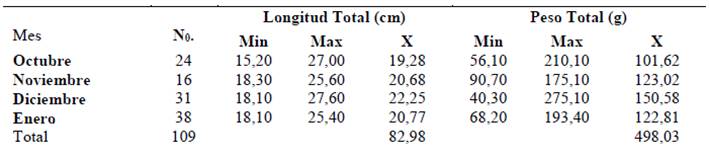
Tabla 2 Distribución de longitud y peso de organismos machos Haemulopsis axillaris, obtenidos en el sector La Caleta, cantón de La Libertad, entre octubre 2017 y enero 2018.
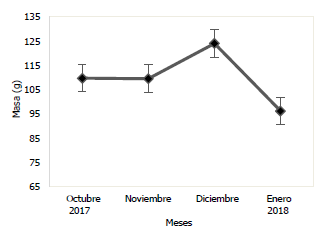
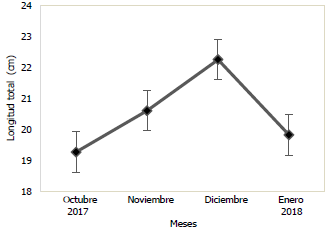
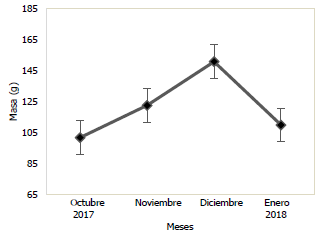
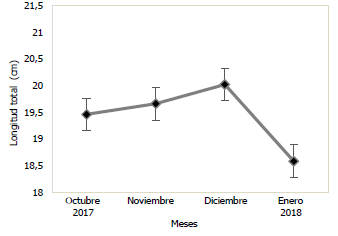
En las Figs. 3 y 4, se puede observar la variación mensual del incremento promedio de la longitud y masa de la población de Haemulopsis axillaris, en la cual se puede apreciar que los más altos valores se registraron en el mes de diciembre 2017, mientras que, en enero 2018, se obtuvieron los valores más bajos. Para las hembras los valores máximos de longitud y masa se obtuvieron en el mes de diciembre 2017, mientras los valores mínimos se registraron en el mes de octubre 2017 (Fig. 5 y 6). Los organismos machos registraron valores máximos de longitud en el mes de diciembre 2017, mientras que existió una variación en los valores máximos de masa, siendo estos registrados en octubre 2017, paralelamente a esto los valores mínimos tanto para longitud como para masa fueron registrados en el mes de enero 2018 (Fig. 7 y 8).
Revista Científica y Tecnológica UPSE Vol. 5, Nº 2 (2018), 10-21
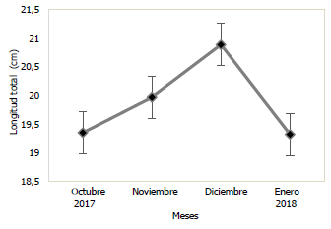
Fig. 3 Variación mensual de la longitud total en ejemplares de la población de Haemulopsis axillaris.
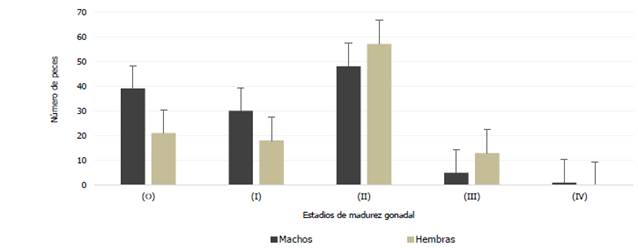
Maduración gonadal hembras
El análisis mensual de los estadios de maduración gonadal de hembras Haemulopsis axillaris indica que los organismos juveniles correspondientes a los estadios 0 a I, fue mayor en los meses de octubre 2017 y enero 2018. Los ejemplares en proceso de maduración, estadio II, se observaron en diciembre 2017 y enero 2018, mientras que la mayor cantidad de organismos sexualmente maduros, estadio III, se observaron en diciembre 2017 (Fig. 9).
Virginal (0)
El 9% del total de organismos examinados (21 individuos) se encontraban en este estadio de maduración gonadal, con promedio de longitud de 17,63 cm y peso promedio de 78,63 g. Los ovarios se presentaron laminares de un color naranja opaco, redondeados en el extremo anterior, registrando valores promedio de longitud 4,10 cm, diámetro 1,30 cm y peso 0,95 g (Fig. 10 A).
Reposo (I)
Constituyeron el 8 % de la población (18 individuos), el promedio de longitud registrado de estos peces fue de 19,16 cm con promedio de peso de 96,13 g. Los ovarios es este estadio se presentaron con un color naranja claro, una longitud promedio de 4,50 cm y grosor promedio de 2,10 cm y además presentaron un peso promedio de 2,10 g (Fig. 10 B).
En maduración (II)
Los organismos en este estadio representaron la mayor parte de la población estudiada con un 24% (57 individuos). La longitud promedio que abarcaron estos peces fue de 21,04 cm con promedio de peso de 127,84 g. Los ovarios presentaron un color naranja más oscuro, vasos sanguíneos marcados, una longitud promedio de 5,30 cm, diámetro de 2,70 cm, peso promedio de 6,55 g (Fig. 10 C) y óvulos translucidos (Fig. 10D).
Maduro (III)
El 6% de los organismos (13 individuos) se encontraba en este estadio gonadal, la longitud promedio fue de 24,80 cm y el peso promedio de 198,07 g. Los ovarios se presentaron abultados ocupando la mayor parte de la cavidad celomática, con un color rosado grisáceo, vasos sanguíneos muy marcados, longitud promedio de 7,40 cm; diámetro promedio de 4,10 cm y un peso medio de 13,30 g (Fig. 10 E).
Se resalta que no se registraron organismos hembras Haemulopsis axillaris en los estadios gonadales correspondiente a Desovante (IV) y Recuperación (V).
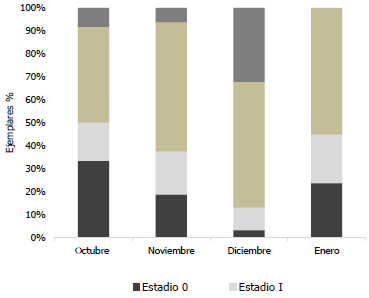
Fig. 9 Proporción de los estadios de madurez sexual mensual de organismos hembra Haemulopsis axillaris

Maduración gonadal machos
El análisis mensual de los estadios de madurez gonadal de machos Haemulopsis axillaris indica que los organismos en proceso de desarrollo sexual (estadio 0 a I) abundaron en diciembre 2017 y enero 2018. Los ejemplares en proceso de maduración (estadio II) se observaron noviembre y diciembre 2017, mientras que la mayor cantidad de organismos sexualmente maduros (estadio III) se observó en diciembre 2017, a su vez el único individuo en periodo reproductivo estudiado fue registrado en el mes de enero (Fig.11).
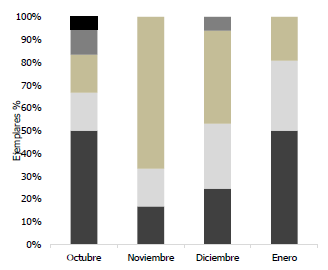
Fig. 11 Proporción de los estadios de madurez sexual mensual de organismos macho Haemulopsis axillaris
Virginal (0)
Representaron el 17 % de la población (39 individuos), el promedio de longitud de los peces fue de 17,09 cm con promedio de peso de 69,87 g. Los testículos presentaron un color blanco translúcido, lóbulos alargados en forma de cinta, longitud promedio de 4,10 cm, diámetro promedio de 0,90 cm y un peso medio de 0,75 g (Fig. 12 A).
Reposo(I)
Constituyeron el 13% del total de organismos estudiados (30 individuos) el promedio de longitud de los peces fue de 18,05 cm con promedio de peso de 94,17 g. Los testículos presentaron una coloración crema, longitud promedio de 4,6 cm, un aumento significativo de grosor con promedio de 2,20 cm y un peso medio de 1,50 g (Fig. 12 B).
En maduración (II)
21% de la población estudiada (48 individuos). La longitud promedio que abarcaron estos peces fue de 20,71 cm con promedio de peso de 123,12 g. Testículos con mayor volumen, tamaño promedio de 5,60 cm, diámetro promedio de 2,60 cm, coloración crema y un peso promedio de 4,10 g. (Fig. 12 C). A su vez, el 45% de la población (machos y hembras) se encontraron en este estadio gonadal.
Maduro (III)
2% del total de organismos estudiados (5 individuos). Estos peces presentaron un promedio de longitud de 25,80 cm. y peso promedio de 192,10 g. T (Fig. 2 D). Testículos de longitud promedio de 7,10 cm, diámetro de 3,8 cm, un peso medio de 5,80 g y una coloración crema (Fig. 12 D).
Expulsante (IV)
Se encontró un solo individuo en este estadío gonadal, con longitud de 35,70 cm. y peso de 539,20 cm. Testículos color crema, abultados que ocupan a mayor parte de la cavidad celomática, longitud 9,70 cm, diámetro de 4,50 cm y un peso de 9,30 g (Fig. 12 E).
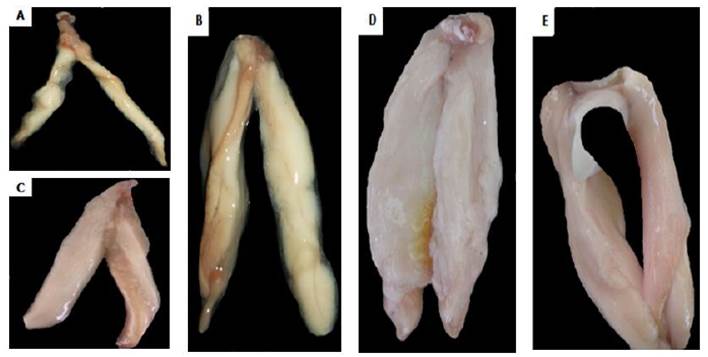
Fig. 12 Estadios de madurez sexual de testículos de Haemulopsis axillaris. A) Estadio virginal (0), B) Estadio reposo (I), C) Estadio en maduración (II), D) Estadio maduro (III), E) Estadio Expulsante (IV) (Microfotografía de estereomicrocospio).
Además, no se registraron organismos machos Haemulopsis axillaris en estadío Post-expulsante (V).
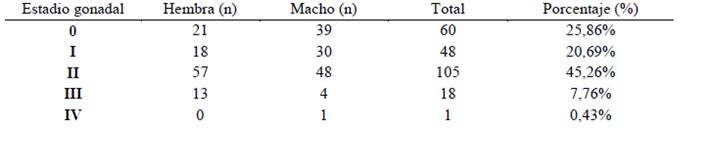
La población general estudiada de Haemulopsis axillaris presentó un predominio del estadío gonadal II, correspondiente a un período de maduración y desarrollo reproductivo, presentando un porcentaje de 42,26%, constituido principalmente por organismos hembras (57 peces); seguido del estadio 0 con un estimado porcentual de 25,86%, evidenciado por peces juveniles en proceso de crecimiento y desarrollo fisiológico, con mayoría de individuos machos (39 peces). Paralelamente el estadio I, se presentó en un 20,69%, con dominio de organismos machos (30 peces); los organismos sexualmente maduros constituyeron el 7,76% de la población, registrando la mayor frecuencia en organismos hembra con alrededor de 13 peces, a su vez el único individuo registrado en estadio IV representó el 0,43% (Tabla 3 y Fig. 13).
Tabla 3. Distribución de número y porcentaje de organismos machos y hembras acorde al estadio de maduración gonadal de la población de Haemulopsis axillaris estudiada
Hembras
Tabla 3 Distribución de número y porcentaje de organismos machos y hembras acorde al estadio de maduración gonadal de la población de Haemulopsis axillaris estudiada
Tamaño de otolitos y edad de organismos acorde a su estadio gonadal
Se extrajeron un total de 463 otolitos sagitta, correspondientes a 231 pares completos (derecho e izquierdo) y un par incompleto (faltante el izquierdo), paralelamente se registraron 4 otolitos con pequeñas fisuras.
La lectura de los anillos de crecimiento en otolitos sagitta, permitió establecer tres grupos de edades (0+, 1+, 2+) (Fig. 14), con variaciones significativas en cada estadio gonadal estudiado.
Estadío 0
El largo total del otolito izquierdo de los organismos en el estadio Virginal osciló entre 0,60 y 0,93 cm, con una media de 0,81 cm, mientras que el ancho osciló entre 0,50 y 0,65 cm, con una media de 0,55 cm. Para el otolito derecho se registraron datos del largo entre 0,45 y 0,95 cm, con una media de 0,81 cm; mientras que para el ancho los valores se encontraron entre 0,45 y 0,77 cm, con un valor promedio de 0,56 cm.
El dominio de edad para este estadio gonadal fue de 0+ (meses), en el 100% de los individuos, observándose la carencia total de anillos concéntricos y la presencia tanto en la cara interior como exterior de estrías radiales orientadas hacia el centro de cada otolito (Fig. 15 A).
Estadío I
El otolito izquierdo mostró valores de largo entre 0,81 y 0,95 cm, con una media de 0,85 cm, mientras que el ancho osciló entre 0,51 y 0,79 cm, con una media de 0,62 cm. El otolito derecho registró valores del largo entre 0,81 y 0,95 cm, con una media de 0,86 cm; mientras que para el ancho los valores se encontraron entre 0,51 y 0,78 cm, con un valor promedio de 0,61 cm.
La edad definida para este grupo de organismos fue de 0+ (meses) en el 100% de los peces, producto de la carencia de anillos concéntricos formados en su totalidad, observándose la presencia de anillos falsos y el predominio de estrías radiales orientadas hacia el centro de cada otolito.
Estadío II
Se registraron valores entre 0,96 y 1,40 cm para el largo del otolito izquierdo y una media de 0,95 cm, mientras que para el ancho los valores oscilaron entre 0,60 y 0,79 cm, con una media de 0,67 cm. Para el otolito derecho se registraron datos del largo entre 0,98 y 1,30 cm, con una media de 0,98 cm; mientras que para el ancho los valores se oscilaban entre 0,59 y 0,80 cm, con un valor promedio de 0,67 cm. La edad dominante para este estadio gonadal fue de 0+ (meses) con un 95% del total de peces, dominando otolitos con presencia de anillos falsos y estrías radiales y en solo el 5% se observó ya la presencia de un anillo concéntrico completo (Fig. 15 B).
Estadío III
El otolito izquierdo mostró valores de largo entre 0,94 y 1,35 cm, con una media de 1,06 cm, mientras que el ancho osciló entre 0,70 y 0,75 cm, con una media de 0,72 cm. El otolito derecho registró valores del largo entre 0,95 y 1,45 cm, con una media de 1,18 cm; mientras que para el ancho los valores se encontraron entre 0,59 y 0,80 cm, con un valor promedio de 0,61 cm.
La edad dominante para este grupo de peces fue de 1+ (1 año) en el 86% de los organismos, observándose la presencia de un anillo concéntrico formado en su totalidad, además de varios anillos falsos y la presencia de estrías radiales orientadas hacia el centro de cada otolito (Fig. 15 C).
Estadío IV
El único individuo (macho) registrado con este estadio gonadal, presentó el otolito izquierdo con un largo de 1,26 cm y un ancho de 0,87 cm; mientras que el otolito derecho presentó un largo de 1,27 cm y ancho de 0,90 cm. Cada otolito presentó dos anillos concéntricos y varias estrías (Fig. 15 D).
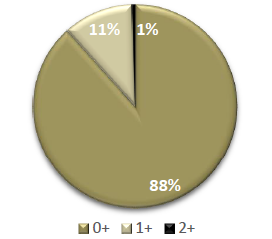
Fig. 14 Gráfica en pastel representando los porcentajes obtenidos en el estudio para los grupos de edad de 0+ (meses), 1+ (1 año) y 2+ (2 años).
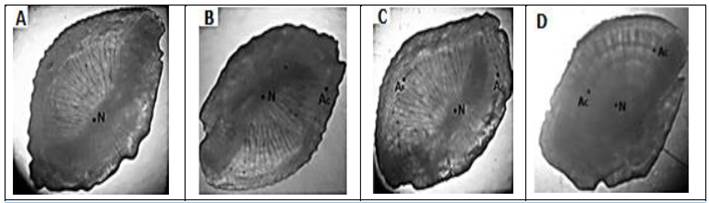
Fig. 15 Anillos concéntricos de otolitos sagitta. A) Otolito sagitta derecho sin presencia de anillos concéntricos B) Otolito sagitta derecho presencia de formación de anillos concéntricos, C) Otolito sagitta izquierdo, presencia de un anillo concéntrico (Ac), anillo falso (Af), D) Otolito sagitta izquierdo, presencia dos anillos concéntricos (Ac).
El análisis de regresión establecido en base a las tallas promedio de 19,38 cm; 24,95 cm y 35,70 cm correspondientes a los grupos de edad 0+ , 1+, 2+ ; permitió evidenciar un coeficiente de correlación de 0,97, demostrando la relación positiva entre la longitud del individuo y la edad de cada organismo.
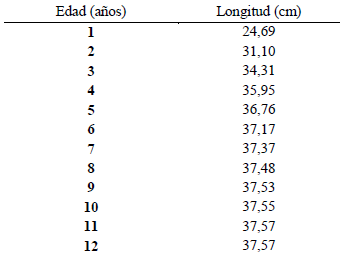
Por otra parte, la correlación longitud-edad, se realizó en base a modelos matemáticos como el método de Taylorel cual permitió establecer una longitud asintótica ( 𝐿∞) de 37,58 cm; mientras que el método de vonBertalanffy estableció un coeficiente de crecimiento (K) de 0,69 𝑎ñ𝑜 −1 y un parámetro de condición inicial (𝑡0) de -0,54. Estos valores reflejan que Haemulopsis axillaris es una especie de crecimiento rápido en los primeros años de vida y alcanza en pocos años su longitud máxima teórica (Fig. 16 y Tabla 5).
Discusión
De acuerdo a las tallas registradas en la investigación, la longitud máxima de Haemulopsis axillaris fue de 35,70 cm, contrario a los señalado por Steindachner (1869) quien indica que la longitud máxima de esta especie es de 30 cm.
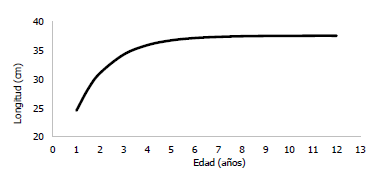
Fig. 16 Curva de crecimiento en longitud y estimación de años, de la especie Haemulopsis axillaris. Fuente: Propia autoría, 2018.
Tabla 5: Estimación de la edad teórica a partir de los valores de los parámetros de crecimiento en longitud de von Bertalanfy y la relación longitud-edad de Haemulopsis axillaris.
En la proporción total de sexo se observa un marcado dominio de los organismos machos, esto se debe a una posible conducta de permanecer mayor tiempo cerca de la superficie a espera de una nueva pareja reproductiva (Duponchelle y Panfili, 1998), contrario a lo mencionado por From (1986) expone que los peces jóvenes son preferentemente de litorales, mientras que los adultos se encuentran mar adentro, es por eso que son capturados muy fácilmente ya que su hábitat no es muy profundo, caso contrario como indica el estudio de Romero (2002) en la plataforma continental de Jalisco y Colima, los organismos pueden tener distribuciones tridimensionales presentando fases estuarinas o con afinidad a otro tipo de fondo de ambientes rocosos cuyos sedimentos proporcionan gran variedad de recursos alimenticios, por eso son fácilmente capturados.
Los organismos hembras sexualmente maduros pertenecientes al estadio III, se observaron en el mes de diciembre; Rodríguez (1985) en Haemulon steindachneri del oriente de Venezuela, y García (1992) en Haemulon plumieri, Haemulon sciurus y Haemulon album de las costas de Cuba; indican que el periodo reproductor de las hembras maduras de estas especies ocurre de marzo a julio, con un periodo de desove que se prolonga hasta el mes de noviembre.
Los parámetros de crecimiento obtenidos por el método de Taylor y método de von Bertalanffy, establecieron que Haemulopsis axillaris es una especie de crecimiento rápido en los primeros años de vida y alcanza en pocos años su longitud máxima teórica debido a que los valores alcanzados de L∞= 37,58 cm y K= 0,69 año-1, valores muy cercanos a la especie Haemulon steindachneri, estudiado en el suroeste de la isla de Margarita (Venezuela), la cual presentó valores con el método de los otolitos de 𝐿∞= 23,10 cm y K=0,569 año-1 (Morales y González, 2009).
Además, el valor alto de K de la especie Haemulopsis axillaris refleja que estos organismos son de vida corta en comparación a otras especies como Dactylopterus volitans con K=0,24 (Longart et al., 2015). No obstante, estos parámetros están relacionado con la tasa metabólica y la mortalidad natural de la población, un 𝐿∞ y K elevados son un indicador de que la población posee altos índices de mortalidad natural (Barreto, 1996).
El recuento de los anillos de crecimiento en otolitos, permitió establecer tres grupos de edad (0+1+, 2+); resultado que contrasta con Morales y González (2009), quienes determinaron en Haemulon steindachneri, (estudiado en la isla de Margarita) cuatro grupos de edad (1+, 2+,3+,4+); resaltando que no se obtuvieron ejemplares pertenecientes al grupo de edad 0+ por no encontrarse en las muestras provenientes de las capturas comerciales de la flota artesanal de Boca del Río (Morales y González, 2009); estas diferencias sugieren la necesidad de establecer medidas de control de captura y pesquerías en el sector La Caleta y el resto de la región donde desembarca este organismo, para evitar la sobreexplotación del recurso, más aún cuando esta especie no posee ningún tipo de regulación pesquera (González et al. 2006).
En la curva de crecimiento en longitud y edad de Haemulopsis axillaris, se evidenció un crecimiento rápido en los dos primeros años de vida, el cual se va reduciendo posterior al segundo año, hasta que alcanzan cerca del cuarto año de vida su longitud máxima asintótica; ante esto Gómez y Larrañeta (1967), indican que existen factores ambientales importantes que influyen sobre la curva de crecimiento, tales como la disponibilidad de alimento y la temperatura; factores que variaran de forma negativa o positiva en el desarrollo del organismo.
Conclusiones
En maduración (II) fue el estadio gonadal predominante tanto en machos y hembras, representando el 45,26% de la población estudiada, seguido del estadio Virginal (0) con un 25,86%, lo cual hace referencia a que los peces de esta especie, están siendo desembarcados en la caleta pesquera de La Libertad en maduración reproductiva inadecuada, al presentar gónadas pequeñas y con gametos en período de desarrollo.
Los organismos que se encontraban en un estadio gonadal III, poseían en su mayoría otolitos con un anillo concéntrico (1 año de edad); los otolitos izquierdos establecieron un promedio de 1,06 cm de largo y 0,72 cm de largo; mientras que la media establecida para el otolito derecho fue de 1,18 cm de largo y 0,61 cm de ancho; estableciéndose de esta forma una diferencia de tamaños entre cada par de otolito, donde se muestra que la estructura ósea derecha tiende a ser más grande y la izquierda a ser más ancha.
La edad dominante para el estadio III fue de 1+ (1 año) con el 86% de los organismos, observándose la presencia de un anillo concéntrico formado en su totalidad, además de varios anillos falsos y la presencia de estrías radiales orientadas hacia el centro de cada otolito.
La lectura de los anillos concéntricos de Haemulopsis axillaris en este estudio, permitió establecer tres grupos de edad 0+, 1+𝑦 2+, respectivamente, con un amplio dominio de 0+, lo cual constituye un indicador de que estos organismos no estas siendo capturados a una edad optima de desarrollo.
La curva de crecimiento establecida en base a la correlación longitud-edad, permitió concluir que esta especie posee un crecimiento rápido durante sus primeros años, son de vida corta y alcanzan en poco tiempo su longitud máxima teórica.

