











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Forma sugerida de citar:
Jiménez Ruíz, E. R., Fonseca González, W. y Pazmiño Pesantez, L. (2019). Patrón de crecimiento durante la primera gestación de vaquillonas cruza con padre Brahman y diferente genotipo materno. La Granja: Revista de Ciencias de la Vida. Vol. 30(2):40-47. http://doi.org/10.17163/lgr.n30.2019.04.
1 Introducción
Calderón et al. (1993); Abeygunawardena y Dematawewa (2004), informaron que las hembras provenientes de cruzamientos entre razas cebuinas y europeas, inician la pubertad a una edad más temprana que las razas puras en condiciones de trópico húmedo, con valores de 15 y 19 meses, respectivamente. La variabilidad genética existente tanto dentro como entre razas se traduce en diferencias en la edad y el peso corporal a la presentación del primer, afectando posteriores a eventos de carácter reproductivo que terminan siendo determinantes en la aptitud productiva global (Navarrete et al., 1995; Nogueira, 2004). Por esta razón, es importante contar con grupos raciales adaptados a estas condiciones particulares, que sean capaces de superar las limitantes productivas que presentan las razas de Bos taurus en estas regiones. En este sentido, se ha observado que las vacas producto de cruzamientos Brahman x Bos taurus presentan mayor fertilidad, mayor producción de leche y mayor longevidad que aquellas de genotipo Bos taurus puro (Grajales et al., 2006; Zambrano-Sepúlveda, 2014). Sin embargo, al aumentar la proporción de genes de origen índico, las hembras tienden a retrasar la edad a la pubertad y ello impacta negativamente en su comportamiento reproductivo posterior. En este sentido, se ha constatado (Rocha y Lobato 2002; Zambrano y Contreras, 2014) la incapacidad de las vacas cruza Brahman x Bos taurus para producir una hembra de reemplazo que mantenga sus mismas características de productividad como consecuencia de la pérdida de la heterosis con respecto a la expresada por la F1.
El registro del peso vivo de los bovinos es una estrategia que permite monitorear el comportamiento en su entorno natural. Esta información se utiliza con diversos propósitos, entre los cuales cabe mencionar la determinación del nivel de alimentación adecuado y el estado nutricional de los animales, el control de la tasa de crecimiento y las respuestas a la selección genética (Marulanda, 1996; Lesosky et al., 2013; Lukuyu et al., 2016).
En resumen, la evaluación del peso vivo de los animales en los sistemas de producción ganadera resulta fundamental si se desean implementar prácticas adecuadas de manejo de los pastizales tendientes al logro de una producción sustentable en la Amazonía. En este contexto, el objetivo del presente trabajo fue evaluar el patrón de crecimiento durante la primera gestación de vaquillonas cruza con padre Brahman y diferente genotipo materno en la Amazonía Ecuatoriana
2 Materiales y Métodos
Se utilizaron datos retrospectivos de 2304 registros individuales del peso corporal de 48 vacas cruzas lecheras pertenecientes a cuatro genotipos: Brahman x Gyr (Gyr) (n=12), Brahman x Brown Swiss (BS) (n=12), Brahman x Jersey (J) (n=12) y Brahman x Sahiwal (S) (n=12) pertenecientes al rodeo lechero del Centro de Investigación, Posgrado y Conservación de la Biodiversidad Amazónica (CIPCA), ubicado en el cantón Carlos Julio Arosemena Tola de la provincia de Napo (Ecuador), en el kilómetro 44 vía Puyo-Tena (coordenadas: S 01° 14.325´; W077° 53.134´) y dispone de una superficie de 42 ha de pastos destinada a la recría. Las vaquillonas evaluadas provenían del mismo establecimiento y habían sido criadas bajo las mismas condiciones ambientales, nutricionales y de manejo e ingresaron al CIPCA con 15-17 meses de edad y pesos corporales (promedio ± EE) de 204 ± 7,7 kg (G); 276 ± 11,0 kg (BS); 204 ± 8,7 kg (J) y 186 ± 6,0 kg (S). A los fines de evaluar el comportamiento dinámico del peso corporal, todos los animales se pesaron cada 30 días en forma individual, entre los meses de diciembre de 2012 y diciembre de 2016. La alimentación del hato bovino en estudio fue de pastoreo libre, con pastizales en base de Brachiaria decumbens (17 585 kg MS/ha/año, Proteína: 10,6% Fósforo: 0,18%; DIV: 44,4%), Brachiaria brizantha (26 970 kg MS/ha/año; Proteína: 10,1%; Fósforo: 0,18 %; DIV: 44,1%), Arachis pintoi (6 212 kg MS/ha/año; Proteína: 19,4%; Fósforo: 0,21%; DIV: 59,2%), Desmodium ovalifolium (5 890 kg MS/ha/año; Proteína: 16,3%; Fósforo: 0,16%; DIV: 39,6%) y Stylosanthes guianensis (15 237 kg MS/ha/año; Proteína: 21,4%; Fósforo: 0,4%; DIV: 48,7%) (Leonard, 2015).
El manejo sanitario aplicado fue el habitualmente empleado para el rodeo bovino del CIPCA, el cual incluye desparasitaciones, baños contra garrapatas y moscas, vacunaciones para fiebre aftosa, rabia bovina y estomatitis vesicular y la aplicación inyectable de vitaminas y minerales. Se registró en forma individual el peso corporal (kg) a la primera preñez, la edad (días) a la primera preñez y el peso corporal (kg) a la primera parición. La normalidad de la distribución de cada una de dichas variables se evaluó con el test ómnibus de D'Agostino y Pearson (D´Agostino y Pearson, 1973), y la homogeneidad de sus variancias con la prueba de Brown-Forsythe (Brown y Forsythe, 1974). Para cada genotipo se calculó el peso corporal promedio a intervalos mensuales durante la primera gestación. Los datos promedio se graficaron en función de los meses de gestación identificándose en todos los casos un comportamiento asimilable a un modelo lineal, el que se constató con un test de rachas o ciclos. El efecto del grupo genético sobre los valores de los estimadores de los parámetros de la función lineal se evaluó con un análisis de la covariancia.
3 Resultados
La Tabla 1 muestra el efecto del grupo genético sobre las tres variables respuesta. Todas ellas mostraron distribución normal y variancias homogéneas. Las diferencias observadas entre genotipos en el peso corporal a la preñez y al primer parto no fueron estadísticamente significativas. Se observaron diferencias en la edad a la primera preñez, correspondiendo la mayor precocidad a la cruza con madre Brown Swiss y la menor precocidad a las cruzas con Gyr y Sahiwal. Las hembras producto del cruzamiento con Jersey presentaron un comportamiento intermedio.
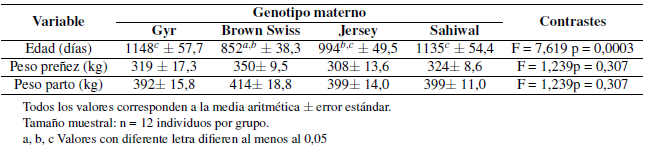
Tabla 1. Edad y peso corporal a la primera preñez en cuatro grupos de vaquillonas F1 con padre Brahman y diferente genotipo materno
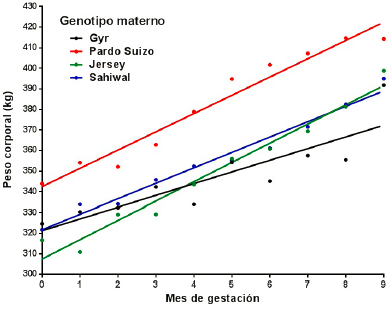
Al ajustar los datos de peso corporal promedio vs meses de gestación se constató una desviación no significativa de la linealidad para los cuatro grupos genéticos [madre Gyr (p = 0,833), madre Brown Swiss (p = 0,283), madre Jersey (p = 0,881) y madre Sahiwal (p = 0,405)]. Los valores del coeficiente de determinación lineal (R2) y las variancias residuales (Sy.x) pusieron de manifiesto una adecuada bondad de los ajustes [madre Gyr (R2 = 0,773; Sy.x = 9,88), madre Brown Swiss (R2 = 0,958; Sy.x = 6,00), madre Jersey (R2 = 0,967; Sy.x = 5,54) y madre Sahiwal (R2= 0,975; Sy.x = 3,85)]. Las ecuaciones de regresión correspondientesa los diferentes grupos se presentanenla Tabla 2.
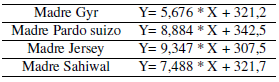
Los valores de la ordenada al origen (estimadores del peso corporal al inicio de la preñez) no mostraron diferencias (p > 0,05) con los valores observados:
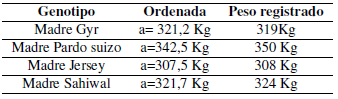
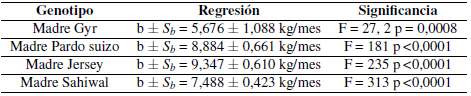
Todas las pendientes fueron estadísticamente diferentes de cero. El análisis de la covariancia puso en evidencia una diferencia estadísticamente significativa en el valor de las pendientes (F = 5,018; p = 0,0058) de los diferentes grupos genéticos, lo que impidió comparar las diferencias en altura. Debido a que las pendientes estiman la tasa de ganancia diaria de peso a lo largo de la gestación, la diferencia observada entre las mismas puso en evidencia un comportamiento particular de las hembras producto del cruzamiento Brahman x Gyr que mostraron un menor aumento de peso en el período. Al excluir a dicho grupo del análisis, las diferencias entre las pendientes de los tres genotipos restantes fueron estadísticamente no significativas (F = 2,845; p = 0,078) lo que permitió calcular una pendiente común a todas ellas (bc = 8,573 kg/mes) y comparar las alturas que resultan estadísticamente diferentes (F = 99,4; p < 0,0001).
4 Discusión
En Ecuador, al igual que en otros países del trópico y el subtrópico de Latinoamérica, ha existido una fuerte tendencia a la sustitución de las razas locales por razas exóticas con el fin de incrementar los niveles de producción de leche. Estas razas, introducidas en la zona con fines de mejoramiento, provienen de regiones donde las condiciones climáticas y nutricionales son generalmente más favorables en comparación con las imperantes en la zona de destino, por lo que su rendimiento productivo en general se ve afectado negativamente. Ante esta evidencia, una estrategia de manejo genético a la cual han recurrido los productores locales es la utilización de cruzamientos entre razas cebuinas (Bos indicus) y razas europeas (Bos taurus), en un intento por minimizar los problemas de adaptación evidentes en estas últimas (Tewolde, 1993; Madalena, 2012).
Pese a sus diferencias de tamaño corporal, todos los genotipos evaluados en este trabajo mostraron similar comportamiento en términos de patrón de crecimiento durante su primera gestación. Este resultado coincide con evidencia previa, según la cual los aumentos de pesos mensuales durante la recría hasta el primer servicio de los cuatro genotipos lecheros cruza analizados fueron de 0,201 kg/día, acordes a la región en estudio. Ninguno de los cruzamientos lecheros se destacó por encima del otro, mostrando todos ellos una paridad en este aspecto (Quinteros et al., 2015).
El hecho que las vaquillonas en estudio no se diferencian en sus ganancias de peso durante la primera gestación, independientemente de su constitución genética, puede interpretarse en términos de una correlación genotipo-ambiente negativa. De acuerdo con ello, al mejor genotipo en términos de velocidad de crecimiento potencial –cruza con Brown Swiss- le corresponde comparativamente el peor ambiente. Esta afirmación se sustenta en el hecho que, si el peso a la preñez representó el 65 % del peso adulto de cada genotipo materno, el peso al parto debería haber sido el 90% del peso adulto, con valores teóricos para Brahman x Gyr de 441 kg, para Brahman x Brown Swiss de 538 kg, para Brahman x Jersey de 473 kg y para Brahman x Sahiwal de 498 kg. Esta predicción difiere de los resultados informados en este trabajo con pesos corporales observados menores a los esperados bajo dicha hipótesis (Tabla 1), y acordes con los que el ambiente limitante de la zona les permitió expresar. La elevada humedad relativa y las altas temperaturas características de la Amazonía ecuatoriana son factores que impactan negativamente sobre las vacas, y de acuerdo a lo planteado por Sánchez (2010); Arias Mader y Escobar, (2008), los animales producirán en la medida que las condiciones del medio ambiente les permitan expresar su potencial productivo. La baja rentabilidad de las producciones refleja los valores resultantes de la evaluación de variables productivas y reproductivas, utilizadas como indicadores de su tasa o nivel de utilidad económica y adaptación de los animales, expresados bajo la forma de promedios de edad al primer parto, intervalo entre partos y presentación celo postparto y producción de leche (Morales, Pérez y Botero, 2009; Motta, Rivera y Mariño, 2012; Quinteros y Marini, 2017). El grupo con genotipo materno Brown Swiss comienza la preñez a una edad más temprana (mayor precocidad), y si bien las diferencias no alcanzaron significado estadístico con mayor peso corporal, esa diferencia se mantiene a lo largo de toda la gestación. Probablemente una mejora en la calidad y en la cantidad de la dieta ofrecida podría aumentar la ganancia media diaria peso; sin embargo, la realidad indica que esta opción no es siempre posible, por lo que se deberían pensar alternativas haciendo uso de los elementos disponibles en la Amazonía. A este respecto, hay antecedentes que demuestran que valores de ganancia de peso comprendidos entre los 600 y 750 gramos por día serían óptimos para las futuras vacas lecheras, mientras que valores menores podrían ser causa de un retraso de la pubertad (Wathes et al., 2014; Gonzáles-Estagnaro et al., 2007). Sin embargo, en general, se trata de trabajos que no han sido realizados en ambientes como el de la Amazonía, por lo que la transferencia de los resultados derivados de ellos no resulta siempre posible.
5 Conclusión
Los resultados permiten corroborar que las condiciones ambientales limitantes reinantes en la Amazonia Ecuatoriana, impiden la expresión de las diferentes potencialidades genéticas de las vaquillonas F1 con padre Brahman y diferente genotipo materno durante su primera gestación, dando como resultado un similar patrón de crecimiento en todas ellas. Teniendo en cuenta que lo que se mide en los animales es un fenotipo que resulta de la acción de su composición genética particular en el medio ambiente en el que le toca expresarse, la elección de una cruza en particular estará determinada por la factibilidad de ofertar un ambiente nutricional más acorde con sus requerimientos. La presencia de una interacción genotipo-ambiente de signo negativo indica que carece de sentido generar por cruzamiento una población cuya potencialidad de crecimiento y su correlato a nivel reproductivo no puede expresarse por limitaciones atribuibles al ambiente.

