











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
I. INTRODUCTION
ROSES, (genus Rosa L.), are enduring woody perennial flowering plants of the Rosaceae family. This genus of roses has an extensive diversity, comprising approximately 200 distinct species and a wide range of 18,000 varieties(1). These plants hold significant importance as ornamental additions, widely cultivated and cherished in parks and gardens across the globe. Beyond their aesthetic value for landscaping, certain species, such asRosa gallicaL.,R. centifoliaL., andR. damascenaMill., have been used for the extraction of rose oil and water(2).
A great number of insect pests attack rose plants at almost every stage and sometimes they cause severe damage to plants(3). The rose sawfly belonging to the family Argidae is a pest that primarily feeds on rose plants and infests different species of roses, includingR. bourboniana, R. multiflora, R. moschata, R. centifolia, R. damascena, andR. indica(4). However, it has been reported on many other host plants such as birch (Betula spp.) including silver birch (Betula pendula) and downy birch (Betula pubescens)(5), Hazel plant (Corylus avellana), Common alder (Alnus glutinosa), Grey alder (Alnus incana), Goat willow (Salix caprea), Grey willow (Salix cinerea)(6), Hawthorn (Crataegus spp.)(7).
The rose sawfly,Arge ochropus(Argidae: Hymenoptera) is a major pest of rose plants and is widely distributed in Asia and Europe(4).Arge paganalarvae mainly feed on rose foliage causing complete defoliation of plants(8). The female rose sawfly lays eggs in newly emerging shoots causing long scratches on the twigs. Rose twigs are often dried out by these scars and the plants ultimately shed their leaves, resulting in reduced yield and quality of roses(9),(10).
Currently, insect pest management primarily relies on the utilization of synthetic insecticides. However, in recent years, these pesticides have not achieved the desired levels of control, possibly due to the development of resistance within the pest populations(11),(12). Moreover, many of the commonly employed pesticides, notably pyrethroids, and organophosphates, pose a high level of toxicity to the natural enemies of insect pests found in the field(13).
Consequently, as a more environmentally sustainable approach to pest control, biological control is frequently explored as an alternative method. Therefore, the utilization of environmentally friendly methods, such as microbial control, holds a significant promise in Integrated Pest Management(14).Metarhizium anisopliae(Metschnikoff) Sorokin -commonly known as the muscadine fungus- holds a significant position in the realm of entomopathogenic fungi (EPF). It has served as a well-established model for the study of biological pest control using fungi(15). Notably, it holds the distinction of being the first fungus to achieve global-scale mass production and deployment for the purpose of insect pest control(16).Metarhizium anisopliae, being a soil-borne entomopathogenic fungus, offers an environmentally sustainable alternative to chemical pesticides(17). Numerous studies have explored the impact ofMetarhizium anisopliaeisolates on various insect pests(18),(19),(20). This approach, utilizing bioinsecticides, is not only effective but also safe for both environmental and agricultural applications(21),(22).
To date, the existing scientific studies addressing sawfly control have predominantly centered around pesticide application and the timing of such applications during egg hatch stages(23),(24). However, as far as our knowledge extends, no study is available on the use of formulated products of EPF to control sawfly larvae.
II. METHODS
A. Insect Rearing
The adults of rose sawfly were collected from the infested rose plants (Rosa indica L.) at the College of Agriculture in the University of Sargodha, and reared in the biocontrol laboratory. The confined pairs of adults were placed in glass cages with nylon nets (45cmX45cmX45cm). Eight small pots with medium-sized rose plants,R. indica(30 cm long) were placed in a rearing cage to provide an innate environment for the female to oviposit. As the female inserted eggs into the tissues of rose twigs, they were moved to separate petri dish. After egg hatching the first instar larvae were placed in clean petri plates. Fresh rose foliage was used as feed for larvae along with sanitation on a daily basis. The culture was maintained in a growth chamber (BIOBASE, BJPX-A400) under controlled conditions of 25± 2°C temperature, 60-70 % RH, and 12:12h (L:D) photoperiod. Pupal cocoons were placed in large glass jars until adult emergence. Cotton soaked with honey solution (30 %) was given to adults as feed. Insects were reared for up to three consecutive generations for further studies.
B. Fungal isolates
The EPF,Metarhizium anisopliaestrain Met F52, (Earth Bio-Sciences in New Haven, CT), was tested against third instar larvae of sawflies. In a controlled laboratory environment, the EPF was cultured on Potato Dextrose Agar (PDA) at conditions of 25 ± 2°C temperature, 70 % relative humidity (RH), and a 12:12h light-to-dark (L:D) photoperiod. The quality of conidia was assessed using a Neubauer chamber hemocytometer. In the subsequent bioassay, a suspension containing 90 % conidial germination was employed, with adjustments made to achieve various concentrations, including 1X106, 1X107, 1X108and 1 X109conidia/ml in distilled water, supplemented with 0.05 % tween 80.
C. Pathogenicity of EPF isolates against A. ochropus
Different concentrations ofMetarhizium anisopliaewere tested against 3rd instar larvae of sawflies taken from the insect culture reared in the laboratory. The treatments were replicated four times and, in each replication, 10 larvae were tested. Forty individuals were released on the fresh rose leaves, kept in petri plates (9mm in diameter) separately, and treated against each concentration. All larvae were treated with different suspensions (1X106, 1X107, 1X108and 1X109conidia ml-1suspension) on the thorax of larvae by using a micro-pipette. The control groups were treated with sterile distilled water with tween 80. After treatment, all petri plates were incubated at 25± 2°C temperature and 60±5 % R.H. Mortality data was recorded on the 7 and 11 days after EPF exposure(25). The effect of each concentration was also recorded on pupal formation.
D. Data analysis
The data were first tested for normality using Shapiro-Wilk test and normalized using log transformation where needed. However, back-transformed means were presented in the results. Data were analyzed by using one-way ANOVA to check the significance ofMetarhizium anisopliaeconcentrations on larval mortality and pupal formation. Means were separated with the least significant difference (LSD) test at a 5 % level of significance. The analyses were performed using Minitab 18.0 software.
III. RESULT
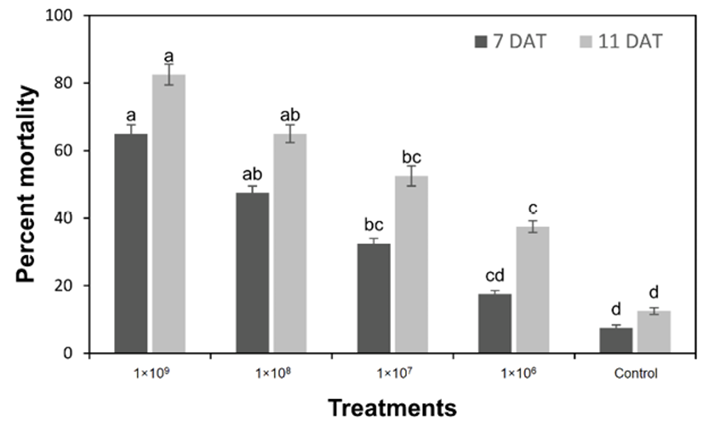
There was a notable effect of different concentrations ofMetarhizium anisopliaeon sawfly larval mortality at 7 days (F = 11.4, P < 0.001) and 11 days (F = 14.5, P < 0.001) of application. The data clearly showed that the mortality of sawfly larvae increased with increasing concentrations ofMetarhizium anisopliae, both after 7 days and 11 days. The highest concentration (1X109conidia/ml) of EPF resulted in the highest larval mortality 82.5 % on the 11 day and 65.0 % on the 7 day, indicating the most effective control of sawfly larvae. With the application of 1X108concentration, the mortality rate was also 65.0 % at 11 days after exposure. Using the lowest concentration (1X106conidia/ml), the larval mortality, was 17.5 % at 7 days and 37.5 % at 11 days of application. In the control group, the mortality rates were much lower at both time points (7.5 % at 7 day and 12.5 % at 11 day) (figure 1).
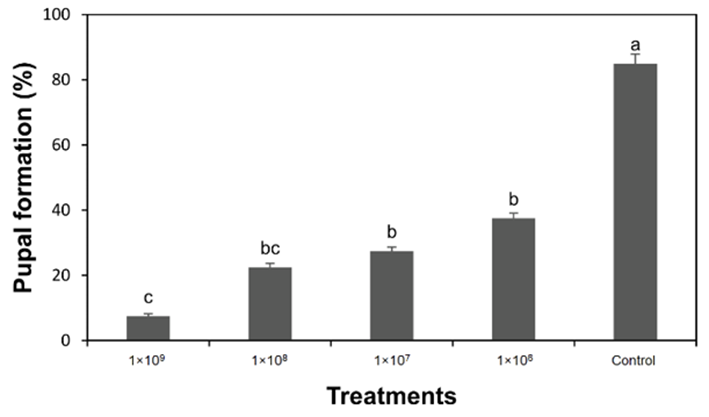
Fig. 1 Percent mortality (means±SE) of sawfly larvae after application of different concentrations ofMetarhizium anisopliaeat 7 and 11 days of applications, means sharing similar letters are not significantly different at P > 0.05.
A significant effect (F = 21.0, P < 0.001) of different concentrations ofMetarhizium anisopliaewas found on pupal formation. The control group, without any EPF treatment, had a high pupal formation rate of 85.0 %, indicating that most larvae in the control group were pupated. In contrast, as the concentration ofMetarhizium anisopliaeincreased, the pupal formation rates decreased. The highest EPF concentration (1X109conidia/ml) resulted in the lowest pupal formation rate at 7.5 %, suggesting that this concentration had the most significant impact on preventing pupation. The pupal formation rate was 37.5 % after the application of a lower concentration (1X106conidia/ml) of EPF.Metarhizium anisopliaeat any concentration inhibited pupation compared to the control (figure 2).
IV. DISCUSSION
Entomopathogenic fungi play a vital role in natural insect population regulation. A wide array of EPF isolates has been extensively examined for their capacity to manage insect pests(26). In this study, we checked the efficacy of different concentrations ofMetarhizium anisopliaein controlling sawfly larvae infesting rose plants. Our results demonstrated a concentration-dependent effect ofMetarhizium anisopliaeon sawfly larval mortality.
The highest concentration tested (1X109conidia/ml) yielded a significant mortality rate, showing the rapid action of the fungus against the larvae. Even the lowest concentration tested had an apparent impact on larval mortality compared to the control group, affirming the potential ofMetarhizium anisopliaeas a sawfly larvae biocontrol agent. Our findings are similar to previous studies related to the successful use of EPFs against rose sawfly. Khosravi et al.(27)evaluated fourB. bassianaisolates (IR-K-40, IRAN403C, SP566, and SPT22) using five distinct conidial concentrations when targeting fourth-instarA. rosaelarvae. Notably, the IRAN403C isolate emerged as the most promising candidate for pest biological control at a concentration of 2X108conidia/mL. Swiergiel et al.(28)reported the potential use of two EPF, a commercial product containingB. bassianastrain GHA and an indigenous isolate (KVL 14-90) ofM. brunneumPetch, for controlling the apple sawfly (Hoplocampa testudineaKlug). Our findings are also similar to Baki et al.(29)who studied the pathogenicity of 17 entomopathogenic fungal (EPF) isolates, encompassing three distinct species:Beauveria bassianawith 14 isolates,Clonostachys rosea(Link) with 2 isolates, andIsaria farinosa(Holmsk) with 1 isolate. The assay results demonstrated a notable and significant enhancement in the bio-efficacy of the tested isolates as time passed, particularly up to 9 days following inoculation.
In addition to larval mortality, we assessed the impact ofMetarhizium anisopliaeon pupal formation. Our results showed a clear inverse relationship between EPF concentration and pupal formation. The control group, compared to EPF treatment, exhibited a high pupal formation, indicating that untreated larvae primarily progressed to the pupal stage under normal conditions. Conversely, the highest EPF concentration (1X109 conidia/ml) led to a low pupal formation rate, emphasizing the potency of the fungus in inhibiting pupation. The trend continued with lower concentrations, demonstrating that even minimal exposure to the fungus can disrupt the normal pupation process. The current study demonstrates thatMetarhizium anisopliaeexhibits pupicidal activity against sawflies. This EPF is capable of breaking down the cuticle layer of pupae, ultimately resulting in their death. Wakil et al.(30)previously documented a reduced pupation rate inRhynchophorus ferrugineus(Olivier) after the application of EPFs. Our findings align with those of Yehia et al.(31), who observed thatB. bassianaandMetarhizium anisopliaealso reduce the pupation percentage of the house fly,Musca domestica. Similarly, Ibrahim et al.(32)reported significant pupal mortality percentages when using EPFs such asB. assiana,Metarhizium anisopliae,andPaecilomyces fumosoroseus.
The method of infection ofMetarhizium anisopliaeon susceptible hosts involves the direct penetration of their cuticle(33). To simplify the explanation of this intricate process, it can be dissected into six distinct stages, as reported by Gao et al.(34):
These stages encompass the attachment of conidia to the host’s cuticle.
The germination and development of conidia.
The formation of specialized structures calledappressoria.
The penetration of the host’s cuticle.
The establishment and colonization within the host’s hemolymph.
The subsequent extrusion and sporulation of the fungus.
Applying the minimal effective dose to control sawflies in rose crops is likely to diminish the infection risk in larvae. However, it’s essential to recognize that the ecological susceptibility in a field setting(35)might vary due to factors like abiotic conditions and local habitat variations. Surviving a fungal infection could have consequences on fitness, such as a potential decrease in lifetime fecundity. Consequently, it is imperative to investigate these potential effects on sawflies.
V. CONCLUSION
The results of this research provide encouraging information regarding the potential application ofMetarhizium anisopliaeas a biological control to combat sawfly larvae infestations in rose plants. Our results highlight the importance of EPF concentration in achieving effective control, with higher concentrations demonstrating superior performance in terms of larval mortality and inhibition of pupal formation. Further research is warranted to explore the practicality and ecological implications of employingMetarhizium anisopliaein field conditions.

