









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El mango (Mangifera indica L.) es un cultivo bien adaptado a las condiciones de Cuba (Capote et al., 1989; Valdés et al., 2015). La antracnosis es la principal enfermedad que ataca este cultivo en el país. La misma es causada por el hongo Colletotrichum gloeosporioides (Penz) Penz & Sacc. fase perfecta Glomerella cingulata, que provoca la caída de la inflorescencia, afecta los frutos, hojas y ramas jóvenes (Álvarez et al., 2006). También aparece como una enfermedad poscosecha de las frutas maduras durante el almacenamiento. Como consecuencia se obtienen malas cosechas con frutos de baja calidad (Rebouca, 2002; Bruwer et al., 2006), lo que conduce a una disminución del precio de la fruta, tanto en el mercado interno como en el internacional (Arauz, 2000; Rodríguez et al., 2002).
El control de las enfermedades producidas por el hongo representa un reto para el agricultor, donde la estrategia de protección preventiva y el factor tiempo son elementos indispensables dado el corto período de incubación del hongo y su alta capacidad de esporulación en tejidos jóvenes (Infoagro, 2006; Abd-Alla y Wafaa, 2010). Por tanto, se hace necesario integrar medidas de manejo como el uso de variedades con cierto grado de tolerancia o resistencia (Carrillo et al., 2005).
En ocasiones implementar medidas de control resulta difícil por la convergencia en un mismo hospedante de varias especies o subespecies de hongos del género Colletotrichum y la gran variedad de formas morfológicas de estas en relación con las variaciones ambientales (Santiago et al., 2005; Butt y Copping, 2000). Para intentar resolver esta situación los estudios se han focalizado, primero hacia la identificación taxonómica de especies y en segundo lugar a la caracterización de sub-poblaciones de cada especie; resultando imprescindible para la implementación de estrategias de control efectivas (Santiago et al., 2005; Jayasinghe y Fernando, 2009).
El presente trabajo tuvo como objetivo caracterizar morfológica, cultural y patogénicamente las cepas de Colletotrichum sp. que conviven en plantaciones de mango en varias de las provincias occidentales de Cuba.
Materiales y métodos
Obtención de los aislamientos: Se colectaron muestras vegetales con síntomas de antracnosis procedentes del municipio de Alquízar, provincia Habana y de parcelas particulares de los municipios Escambray y Jaruco, ubicados en las provincias Cienfuegos y La Habana respectivamente. Las porciones afectadas se lavaron con agua corriente, se desinfectaron con hipoclorito de sodio al 1% durante 1 min y se sembraron en cajas Petri con medio de cultivo Agar-Papa-Dextrosa (PDA), se incubaron a 30oC hasta que se observó crecimiento micelial. A partir de estas se realizaron cultivos monospóricos mantenidos a 30oC en oscuridad. Una vez crecidos, se conservaron en el cepario en tubos de ensayo a 4oC.
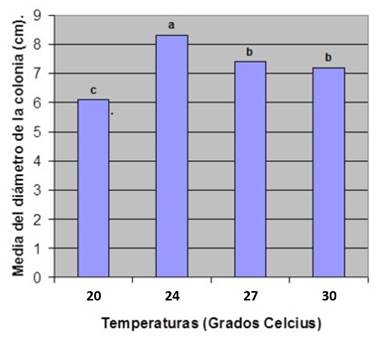
Caracterización cultural de los aislamientos: Los aislamientos del cepario se sembraron en cajas Petri con PDA y se incubaron a 27 ± 2oC, durante 7 días. Una vez crecidas las colonias se tomaron discos con sacabocados de 5 mm de diámetro y se colocaron en el centro de cajas Petri con PDA, de 11 cm de diámetro. Se evaluaron 3 réplicas por aislado a las temperaturas 20, 24, 27 y 30oC, durante 7 días. Pasado este tiempo se realizó la observación de las características culturales y medición de las colonias siguiendo la metodología de Freeman et al. (1998). A los datos del diámetro de la colonia se les realizó un análisis de varianza bifactorial con dos tratamientos y tres réplicas por tratamientos. Las medias fueron comparadas por el Test de Rangos Múltiples de Duncan (Cigarroa, 1985).
Observación de la fase sexual: Los aislados del cepario se sembraron en cajas de 11 cm de diámetro con PDA y se incubaron a 30oC bajo un ciclo de 12 horas luz y 12 horas oscuridad. A partir de estas cajas se realizaron observaciones semanales, por espacio de 2 meses, para determinar la formación de peritecios y ascosporas propios de la fase sexual (Glomerella cingulata).
Determinación de la concentración conidial: Para evaluar la concentración de conidios por aislado, se prepararon suspensiones adicionando 5 discos de micelio, obtenidos a partir de colonias de 15 días de incubación a 30oC, en 10 ml de agua destilada estéril como se describió anteriormente. A partir de estas se determinó la concentración en Cámara de Neubauer. A los datos obtenidos se les realizó un análisis de varianza de clasificación simple (ANOVA) y las medias fueron comparadas por el Test de rangos múltiples de Duncan (Cigarroa, 1985).
Caracterización morfométrica de los aislamientos
Morfometría de los conidios: A partir de colonias con 7 días de incubación a 30oC, se tomaron 5 discos de 5 mm de diámetro por aislado y se adicionaron en 10 ml de agua destilada estéril, colocándose en un vortex durante 1 min para propiciar la liberación de los conidios. La concentración de las suspensiones se ajustó a 1 x 105 conidios.ml-1. A partir de estas se hicieron preparaciones y se midieron 50 conidios por aislado, describiéndose su forma según (Gutiérrez et al., 2001). Esto se realizó con la ayuda del microscopio óptico Olympus con un aumento de 100x.
Morfometría de los apresorios: Se prepararon portaobjetos recubiertos con una película fina de Agar-Agua y se adicionaron 50 µl de suspensiones ajustadas a 1 x 105 conidios.ml-1. Se realizaron tres réplicas por aislado. Los portaobjetos se incubaron a temperatura ambiente durante 24 horas, pasado este período se observaron al microscopio óptico Olympus, con aumentos de 40 y 100x, midiéndose y describiéndose 50 apresorios por cada aislado, según Gutiérrez et al., 2001.
Prueba de patogenicidad: La capacidad infecciosa de los aislamientos se evaluó en hojas de plantas de mango de las variedades Haden, Filipino, Señora y Keitt, procedentes de la colección de mango de la Estación Experimental de Frutales del municipio de Alquízar, provincia Artemisa. Se colectaron 90 hojas jóvenes y sanas de cada variedad. Estas se lavaron con agua, detergente y alcohol al 70 % para eliminar suciedades y desinfectar respectivamente. Luego se colocaron en cajas Petri conteniendo portaobjetos y papel de filtro humedecido con agua estéril.
Se montaron tres réplicas y un testigo por cada variedad y cada aislado, realizando 6 incisiones con una aguja en cada hoja. Como inóculo se usó 10 µl de una dilución de 1 x 105 conidios.ml-1para cada aislado. En el caso de los testigos, se aplicaron 10 µl de agua destilada estéril en lugar de la suspensión. Las cajas se incubaron a temperatura ambiente y se observaron diariamente por espacio de 5 días, siguiendo la aparición de síntomas y midiendo el diámetro de la lesión (virulencia de la cepa). Los datos del diámetro de la lesión por aislado y variedad se procesaron usando la transformación (x+1) y se analizaron siguiendo un análisis de varianza trifactorial. Las medias fueron comparadas por el Test de rangos múltiples de Duncan (Cigarroa, 1985).
Análisis estadístico para la agrupación de los aislados: Se realizó un análisis estadístico de componentes principales (ACP) con los resultados obtenidos para determinar la variable que más aportó a la diversidad. Se hizo un análisis de agrupamiento con las tres variables que más aportan utilizando la distancia euclidiana, basado en la media aritmética no ponderada y el algoritmo SAHN del paquete de programa NTSys versión 2.1
Resultados y discusión
Obtención de aislamientos
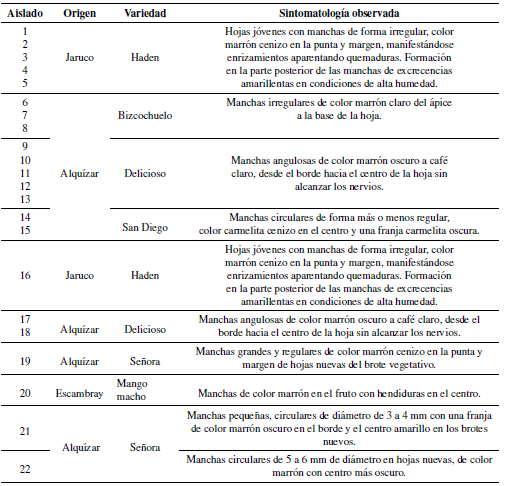
De acuerdo a su origen y la variedad de mango con la que se trabajó se obtuvieron 22 aislamientos fúngicos que aparecen relacionados en la Tabla 1.
Tabla 1. Relación de aislados de Colletotrichum sp. obtenidos de acuerdo a su localización y variedad de plantas de mangos
Caracterización cultural de los aislamientos
En general para las temperaturas de 20, 24, 27 y 30°C el crecimiento fue siempre circular y con bordes regulares excepto para los aislados 9 y 20 que presentaron bordes irregulares. Las coloraciones de las colonias variaron entre el blanco (Figura 1: D y E), gris verdoso (Figura 1: A y B) y gris pálido (Figura 1: C y F).

Figura 1. Diferentes tipos de crecimiento de las colonias de los aislados en caja Petri con PDA a 27± 2oC, después de 7 días incubación durante es estudio
En la mayoría se encontraron sectores de crecimiento (Figura 1: A, C y D) y en muy pocos, halos concéntricos de crecimiento (Figura 1: B y E). Ambos tipos de zonación fueron también comúnmente observadas en el reverso de las colonias y en el caso de los aislamientos 4, 5, 11, 13 y 17 los sectores formaban una estrella. Se manifestaron diferentes tipos de texturas en las colonias: harinosa (Figura 1: F), lanosa (Figura1: A y B), algodonosa (Figura1: C y D) y afelpada (Figura1: E), pudiendo presentarse diferencias de textura del centro al margen de la colonia (Figura 1: A). Solo los aislados 10, 13, 14 y 15 presentaron pigmentos de color rosa salmón. La presencia de exudaciones sobre el micelio se manifestó de forma variable entre los aislados a las diferentes temperaturas por lo que no puede catalogarse como factor distintivo de una cepa o temperatura. Estos resultados coinciden con los reportados por Sangeetha y Rawal en el 2008 quienes obtuvieron aislamientos con semejantes características. La temperatura de 27°C favoreció la esporulación respecto a las restantes. La masa conidial se presentó principalmente como exudaciones de color naranja, viscosas que se desarrollaron a partir de acérvulos maduros sin setas; concordando con lo planteado por Arauz (2000) y Gutiérrez et al. (2001).
Observación de la fase sexual
En general, los aislamientos que formaron fase sexual se observaron pocos peritecios y la mayoría de ellos estaban inmaduros. Solo los aislamientos 2, 3 y 5 los presentaron de color negros y errumpentes en medio de cultivo.
La morfología y dimensión de las ascosporas permite establecer que las cepas periteciales pertenecen a la especie Glomrella cingulata (Stonem) de acuerdo a lo publicado por Freeman et al. (1998); Gutiérrez et al. (2001) y Carrillo et al. (2005). En los aislados 2 y 3 se observaron ascas maduras con ascosporas de 12-20 x 5-6 μm, hialinas, redondeadas, multivacuoladas y curvadas (Cerón et al., 2007).
Determinación de la concentración conidial
Los resultados del conteo de conidios en Cámara de Neubauer aparecen en la Tabla 2. Se encontraron diferencias significativas (P<0.05) entre los aislamientos en el número de conidios producidos. En todos los casos la concentración de conidios se encontró por encima de 1 x 105conidios.ml-1coincidiendo con lo informado por Sanders et al., 2000 y Sangeetha y Rawal (2008). Sobresalen los aislados 4 y 5 con concentraciones del orden de 107 conidios.ml-1.

Caracterización morfométrica de los aislamientos
Morfometría de los conidios
Los aislamientos formaron conidios hialinos, cilíndricos y rectos. Su largo osciló entre los 15.7 y 11.1 μm y su ancho entre los 3.0 y 4.5 μm (Tabla 3). En su mayoría presentaron ambos extremos redondeados aunque un porciento considerable presentó al menos un extremo ahusado lo que coincide con lo encontrado por Sanders et al. (2000) y Sangeetha y Rawal (2008). Todos los conidios presentaron abundantes cuerpos esféricos como publica Kuo (1999). En los aislados 6, 8, 13 y 19 entre el 1 y el 3% de los conidios fueron fusiformes.

Morfometría de los apresorios
Se observaron apresorios lobulados o irregulares, clavados y ovoides, de color pardo oscuro. La forma irregular predominó sobre las restantes como aparece reflejado en la Tabla 3. En todos los casos se observaron apresorios terminales en tubos germinativos con tallas en un rango de 9.7-14.5 x 5.5-10.1 μm. Esto coincide con Páez (1996); Arauz (2000) y Sangeetha y Rawal (2008) quienes observaron apresorios lobulados, pardo oscuros y que podían ser terminales en tubos germinativos o salir directamente de los conidios.
Estas características coinciden con la información registrada en la descripción de la especie Colletotrichum gloeosporiodes (Penz) Penz & Sacc por Sutton (1980), quien define que esta presenta conidios rectos, cilíndricos, de 9.0-24.1 x 3.0-6.7 μm y apresorios de 6-21 x 4-12.7 μm, clavados o irregulares. Por otra parte, Carrillo et al. (2005) plantean que la mayoría de los aislados de Colletotrichum gloeosporioides forman conidios con ápices redondeados y menos del 10% con ápices ahusados.

Figura 3. Pruebas in vitro de la patogenicidad de los 22 aislados y sus correspondientes testigos en hojas de diferentes variedades de mangos. (A-aislado 21 variedad Filipino; B-aislado 7 variedad Señora; C-aislado 13 variedad Filipino; D-aislado 10 variedad Keitt; E-aislado 12 variedad Haden; F-aislado 20 variedad Haden; G-aislado 3 variedad Señora y H-aislado 6 variedad Keitt).
El estudio in vitro de la virulencia de los 22 aislados mostró que las infecciones experimentales fueron positivas, ejemplos ilustrados en la Figura 3. Las hojas mostraron manchas de color marrón, con forma regular, las mismas aumentaron gradualmente y en algunos casos llegaron a cubrir totalmente el órgano de la planta durante el tiempo de estudio. Al cabo de los 5 días, en las lesiones hubo crecimiento micelial emergente y aparecieron acérvulos con abundante producción de conidios inmersos en masas cromógenas, anaranjadas en su mayoría y salmón solo en el caso del aislado 21. En ningún caso se produjeron peritecios propios de la especie Glomrella cingulata (Stonem) Spauld & Schrenk. Los materiales usados como testigos permanecieron siempre libres de infección.
Las lesiones con mayor velocidad de crecimiento para todas las variedades fueron las provocadas por las cepas 21, 19, 3, 2, 17 y 5 en ese orden respectivamente. Dentro de estas se encuentran tanto cepas periteciales como productoras únicamente de conidios, contrastando con los resultados obtenidos por Gutiérrez et al. (2001) quienes plantean que las lesiones causadas por la cepa peritecial, crecen más rápidamente que las ocasionadas por la conidial.
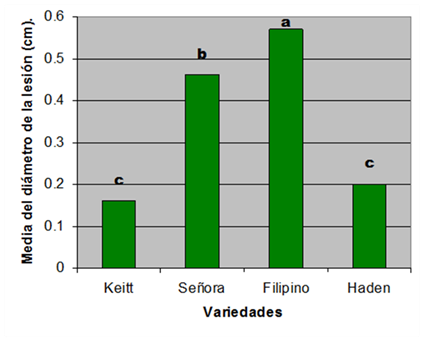
Figura 4. Medida de la sensibilidad de las variedades de mango resultado de las pruebas de patogenicidad de los 22 aislados de Colletotrichum gloeosporiodes (Penz) Penz & Sacc.
Se observaron diferencias significativas (P<0.05) entre las variedades (Figura 4). La variedad Filipino fue la más afectada contraponiéndose a lo planteado por Cañizares (1966) y Capote et al. (1989) quienes la incluyen dentro de las más resistentes a la antracnosis. Estas diferencias respecto a las demás variedades puede deberse a que el la variedad de mango Filipino es considerada por los agricultores como una variedad temprana, que su período de maduración comienza en el mes de abril, quizás esta baja susceptibilidad está referida no a una resistencia genética sino a que los frutos se producen y maduran temprano, fuera de la época de lluvias en que se favorece la infección por el patógeno, por tanto, no contradicen los resultados obtenidos. La variedad Señora mostró una alta susceptibilidad al patógeno coincidiendo con Capote et al. (1989).
Entre las variedades Haden y Keitt no hubo diferencia significativa. Ambas presentaron “in vitro” bajo nivel de susceptibilidad. Estos resultados coinciden con lo planteado por Capote et al. (1989), para quien estas se agrupan dentro de las variedades con baja susceptibilidad a la antracnosis en el campo. Sin embargo, Campbell (1992) y Rodríguez et al. (2002) publican sensibilidad a la enfermedad para la variedad Haden. Estas divergencias pueden tener su base en la influencia de las diferencias climáticas sobre el ciclo de la enfermedad y la maduración del fruto, debido a que los estudios se realizaron en diferentes regiones.
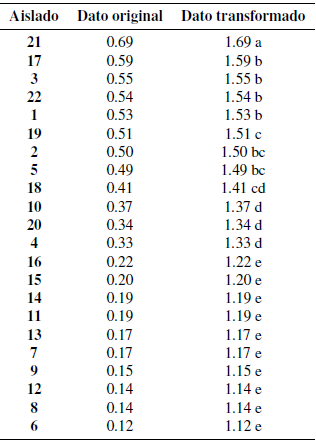
La virulencia de los aislados es presentada en la Tabla 4, donde los menos patogénicos son aquellos que producen menor cantidad de conidios teniendo en cuenta los resultados del conteo realizado. Los aislados obtenidos de Haden y Señora fueron más patogénicos en estas que en el resto de las variedades, concordando con Sangeetha y Rawal (2008) y Denoyes et al. (2003) quienes plantean que las cepas obtenidas de un hospedero particular eran más patogénicas en este que en otros hospederos.
Tabla 4. Relación de agrupamiento de los 22 aislados de acuerdo a su virulencia en las pruebas de patogenicidad con la correspondiente transformación x+1, S = 0.03 (P<0.05)
Se reprodujeron los mismos síntomas observados en las muestras del campo y los reaislamientos mostraron las mismas características que los originales. Según los postulados de Koch, las cepas aisladas fueron las causantes de la antracnosis en las muestras colectadas.
Análisis estadístico para la diferenciación de los grupos
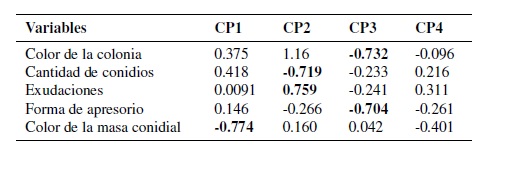
Los factores que más influyen en el agrupamiento de los aislados se corresponden con el color de la colonia, la cantidad de conidios, las exudaciones al medio, la forma del apresorio y el color de la masa conidial como se muestra en la Tabla 5.
Tabla 5. Resultado estadístico en el análisis de componentes principales hasta la cuarta componente, con los aportes de variables cualitativas y cuantitativas.
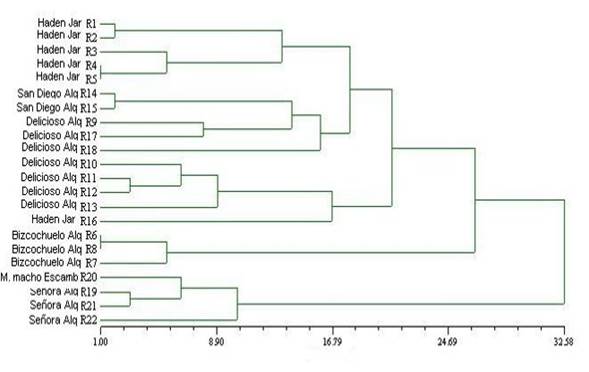
La Figura 5 nos muestra el dendograma de los aislados de C. goleosporioides; estos resultados indican que los aislados se agrupan fundamentalmente atendiendo a la localidad de donde fueron obtenidos en la investigación. En el primer grupo (I) formado por los aislados de Jaruco var Haden que se diferencian perfectamente del grupo (II), quienes en su mayoría están constituidos por los aislados obtenidos a partir de diferentes variedades de mango colectadas en la localidad de Alquilar. Existe un aislamiento procedente del Escambray que se agrupa al final y no se corresponde con ninguno de los grupos mencionados anteriormente. En este sentido coincidimos con Torres (1995), quien planteó que las diferentes regiones climáticas de Cuba determinaban el comportamiento del cultivo, Gutiérrez et al. (2001), en estudios realizados afirmó que existen aislados de C.gloeosporioides que se agrupan más que por su variabilidad por la zona donde se realizó el aislamiento, indicando que el ambiente puede modificar el comportamiento de las cepas.
Conclusiones
Las 22 cepas aisladas en la región occidental de Cuba, pertenecen a la especie Colletotrichum gloeosporioides (Penz.) Penz. and Sacc con una temperatura óptima de crecimiento por encima de 24 ± 2oC. Las dimensiones de las estructuras medidas: conidios y apresorios, se encuentran entre los límites establecidos mundialmente para esta especie. La patogenicidad in vitro de los aislados se manifiesta en concordancia de la variedad de planta, siendo el aislado 21 con más grado de virulencia.