











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Forma sugerida de citar:
Ugas Pérez, M., Calderón Castellanos, R., Rivas Meriño, F., y Núñez Ravelo, F. (2022). Cuantificación del flujo de CO2 en el suelo colonizado por Avicennia germinans, emplazado en el humedal Laguna Grande, sector Los Totumos, estado Miranda, Venezuela. La Granja: Revista de Ciencias de la Vida. Vol. 35(1):85-97. http://doi.org/10.17163/ lgr.n35.2022.07.
1. Introducción
El suelo es un importante reservorio de carbono, ya que en sus primeros metros se pueden hallar alrededor de 1500 Pg C (Petagramos de carbono), mientras que en la atmósfera se estiman unos 800 Pg C, y en la vegetación terrestre aproximadamente unos 500 Pg C. Sin embargo, este se encuentra en constante movimiento en diferentes formas moleculares (Organización de las Naciones Unidas para la Alimentación y la Agricultura, 2017).
El dióxido de carbono (CO2), constituye uno de los derivados del carbono orgánico que se encuentra en permanente interacción en el sistema climático. En la relación suelo-atmósfera, este gas invernadero es producido por diversas condiciones: (a) por la respiración de los micro y macro organismos, así como por las raíces vivas en la rizosfera, además de la materia orgánica descompuesta directamente de las plantas o a través de las cadenas tróficas (Departamento de Agricultura de los Estados Unidos, 1999; Organización de las Naciones Unidas para la Alimentación y la Agricultura, 2017), y (b) absorbido en el suelo desde la atmósfera, por las plantas en el proceso de fotosíntesis, mediante el cual se fija el carbono para producir biomasa y se libera la partícula de oxígeno a la atmósfera (Visconti y De Paz, 2017).
Ahora bien, este secuestro de CO2 en la estructura del suelo es de gran importancia para el ciclo climático global, ya que contribuye con el equilibrio de los gases de efecto invernadero en la atmósfera. Es importante destacar que los bosques cubren el 29% de la superficie emergida del planeta, y el carbono que se encuentran en sus suelos constituye aproximadamente un 36% del total localizado a un metro de profundidad (Zambrano, Franquis e Infante, 2004), siendo la respiración del suelo el flujo más importante del ciclo del carbono (López y Monterroso, 2020), representando aproximadamente el 75% del total en estos ecosistemas (Law y col., 2001). En el caso particular del manglar, Moreno y col. (2002); Kauffman, Donato y Adame (2013), coinciden en referir que estos contienen las reservas más grandes de carbono, debido al desarrollo de un complejo ecosistema que contribuye con la captura del CO2 en el suelo.
De acuerdo con lo referido por Kao, Freyre y Balser (2010), debido a la dominancia de ciertas plantas típicas en ambientes tipo humedales, se produce el flujo de oxigeno desde la atmósfera hacia la zona superficial y subsuperficial del suelo, específicamente hacia la rizosfera, propiciando la producciónoxidación de gases de efecto invernadero, de allí que el aporte y calidad del carbono orgánico puede estar influenciada por la capacidad forestal de la especie dominante. Es posible que por estar ubicados en zonas intermareales, es decir, en la transición entre la zona de marea alta y marea baja, este tipo de bosques se encuentren inundados periódicamente por acción de la pleamar, por lo que el suelo pasa por procesos dinámicos de oxidación-reducción, generando condiciones propicias para la activación de procesos microbiológicos, tales como la nitrificación, desnitrificación y metanogénesis, generadores de gases invernaderos que en ocasiones puede ser liberado a la atmósfera (Chen y col., 2016).
Por su parte Sánchez y col. (2011), refirieren que los suelos de los humedales donde se desarrollan los manglares almacenan por largo tiempo el carbono, motivado a que estos por lo general se mantienen mayormente inundados. No obstante, debido al poco reconocimiento de la importancia de estos reservorios, aunado a los impactos generados por las acciones antropogénicas, estos se han reducido en un 35% del total. En efecto, la acción antropogénica asociada con los diversos usos del suelo, aunado a la tala y la quema de grandes extensiones de terreno, han generado perturbación de los ecosistemas que constituyen reservorios, y en consecuencia un desequilibrio ambiental, propiciando que los suelos pasen de ser sumideros a fuentes de emanación de CO2, potenciando el calentamiento global en el planeta (Sánchez y col., 2011; Visconti y De Paz, 2017; Organización de las Naciones Unidas para la Alimentación y la Agricultura, 2017).
De allí que Villalobos (2012), expresan que es necesario gestionar de la manera más apropiada los hábitats que son reservorios de carbono, tal como los manglares, a fin de conocer su capacidad forestal para la producción-almacenamiento de los gases de efecto invernadero, con el objeto de contribuir con la mitigación de los impactos generados al sistema climático. En este sentido, se han llevado a cabo investigaciones que permiten conocer las potencialidades forestales de los bosques de manglar, referentes a la producción y secuestro de CO2; entre estas, los aportes de Kauffman, Donato y Adame (2013) y Herrera y col. (2016), quienes expresan que México y Brasil forman parte de los 4 países con mayor reserva de manglares a nivel mundial. Por su parte, Moreno y col. (2002), Lozano (2007) y Sánchez y col. (2011), coinciden que, dentro de los humedales costeros, los manglares son un reservorio importante para la fijación o captura de carbono.
Para Costa Rica y Colombia, Yepes y col. (2016) y Villalobos (2012), respectivamente, afirman que los manglares forman parte de los ecosistemas más productivos del planeta en cuanto a producción de carbono, por lo que constituyen un ambiente clave para la mitigación del calentamiento global. Venezuela, por encontrarse en plena zona intertropical y poseer una extensa línea de costa, presenta importantes comunidades de manglar, sobre los cuales se han llevado a cabo diversos estudios dirigidos hacia el reconocimiento de su distribución geográfica, así como su caracterización estructural con base a los métodos de la biología, botánica y la agronomía, (Pannier y Pannier, 1989; Medina y Barboza, 2003; López, Barreto y Conde, 2011; Cumana, Prieto y Ojeda, 2000; Bonilla y col., 2010; Romero y Meléndez, 2013).
Núñez y col. (2019), reportaron para el manglar ubicado en Boca de Uchire, localizado a 94,93 km al sureste de Los Totumos, diferencias en el flujo de CO2 en suelos no rizosféricos, en contraste con los dominados en superficie por Avicennia germinans y Conocarpus erectus. De igual forma, Sánchez, Paolini y Rodríguez (2010), identificaron variaciones en el flujo de CO2 en suelos colonizados por Rizhophora mangle en la isla de Margarita, estimando que debido a que las condiciones climáticas inciden en la disponibilidad de sal en el suelo, esta afecta el flujo del referido gas. Como se evidencia, los estudios sobre la cuantificación del carbono orgánico, así como los flujos de dióxido de carbono en estos ecosistemas son escasos, y hasta ahora no se ha desarrollado en la zona de estudio: humedal Laguna Grande, sector Los Totumos. Esta zona se encuentra ubicada al suroeste de Cabo Codera, astronómicamente se localiza entre los 10◦ 320 3400 y los 10◦ 320 4400 de latitud norte, y entre los 66◦ 40 4400 y 66◦ 40 5400 de longitud oeste, limitando al norte con la Cordillera de la Costa, al oeste con bahía de Buche y al sur y al este con el Mar Caribe. (Figura 1), se extiende en 7,77 ha, de las cuales el bosque monoescífico de Avicennia germinans ocupa 4,42 ha, lo que representa el 56,88% de la superficie total (Figura 2).
De acuerdo con Garrido (2017), geomorfológicamente el área constituye un depósito tipo playa semielongada con una longitud de aproximadamente 400 metros, y presenta un relieve suave con mayor desarrollo hacia los extremos norte y sur que en la zona central, constituido fundamentalmente por partículas de la talla de la arena media, con predominancia de bioclastos. Los datos de las estaciones climáticas Carenero y Tacarigua Mamporal, adscritas a la Dirección de Hidrología y Meteorología de (Ministerio del Poder Popular para el Ambiente, 2013a; Ministerio del Poder Popular para el Ambiente, 2013b), localizadas en la misma línea de costa sin marcadas diferencias altitudinales ni latitudinales entre estas y Los Totumos, permiten afirmar que el área presenta temperaturas correspondientes a un régimen isotérmico, con una media anual estimada de 26,75 ◦C, siendo los meses más cálidos abril y octubre con 27,7 ◦C, y los meses más frescos de noviembre a marzo, con un mínimo registrado de 25,15 ◦C. Las precipitaciones se distribuyen en un régimen unimodal para alcanzar un monto anual de 1141,3 mm, cuyo periodo lluvioso se extiende desde junio a diciembre. La evaporación es elevada durante todo el año, estimándose un monto anual de 1.781,9 mm (Figura 3).
De acuerdo con la clasificación de Goldbrunner (Foghin, 2002), el área se ubica en el piso térmico tropical, con altitudes que no sobrepasan los 10 msnm. En cuanto a los rasgos edáficos, predomina el orden Entisols, con suelo caracterizado por presentar un material poco desarrollado con profundidad menor a 10 metros, un horizonte superficial incipiente que descansa sobre el material rocoso, textura arenosa, altamente salino y alcalino, del suborden Orthens y del gran grupo de los Torriorthens (Cárdenas, 1965; Elizalde, viloria y Rosales, 2007; Gobernación del Estado Bolivariano de Miranda, 2010).
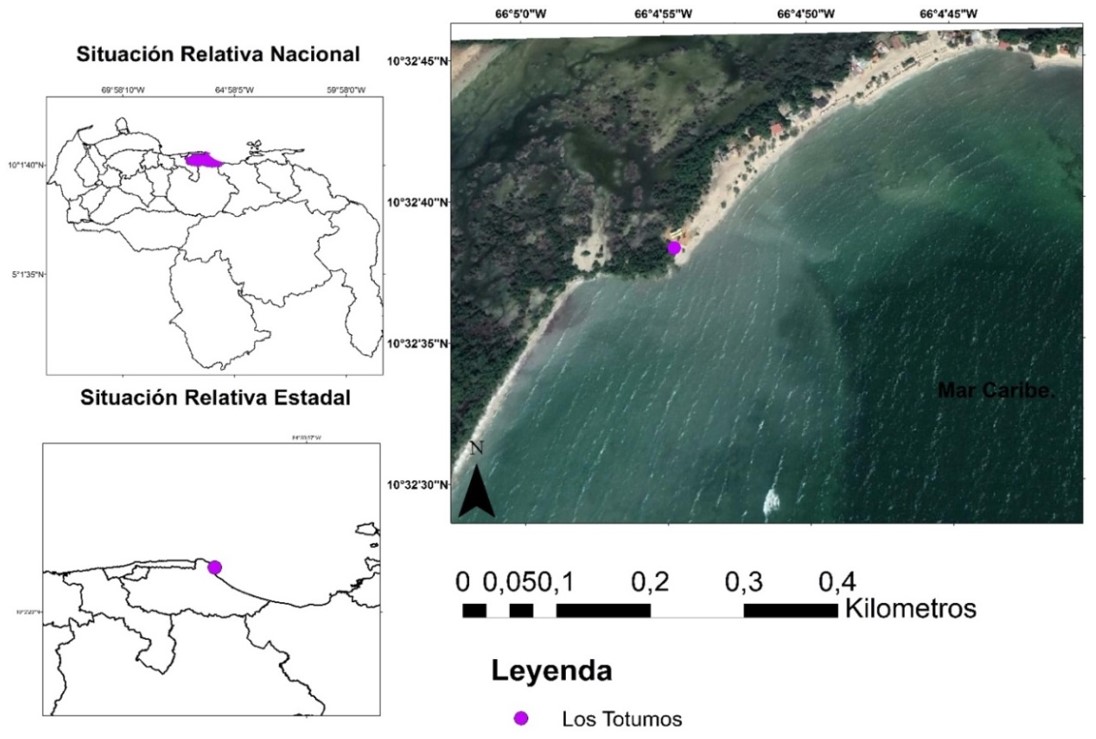
Figura 1. Mapa de Localización de Los Totumos. Elaborado a partir de la Imagen Satelital Lansatd 8. Combinación de las bandas 321: Color natural.
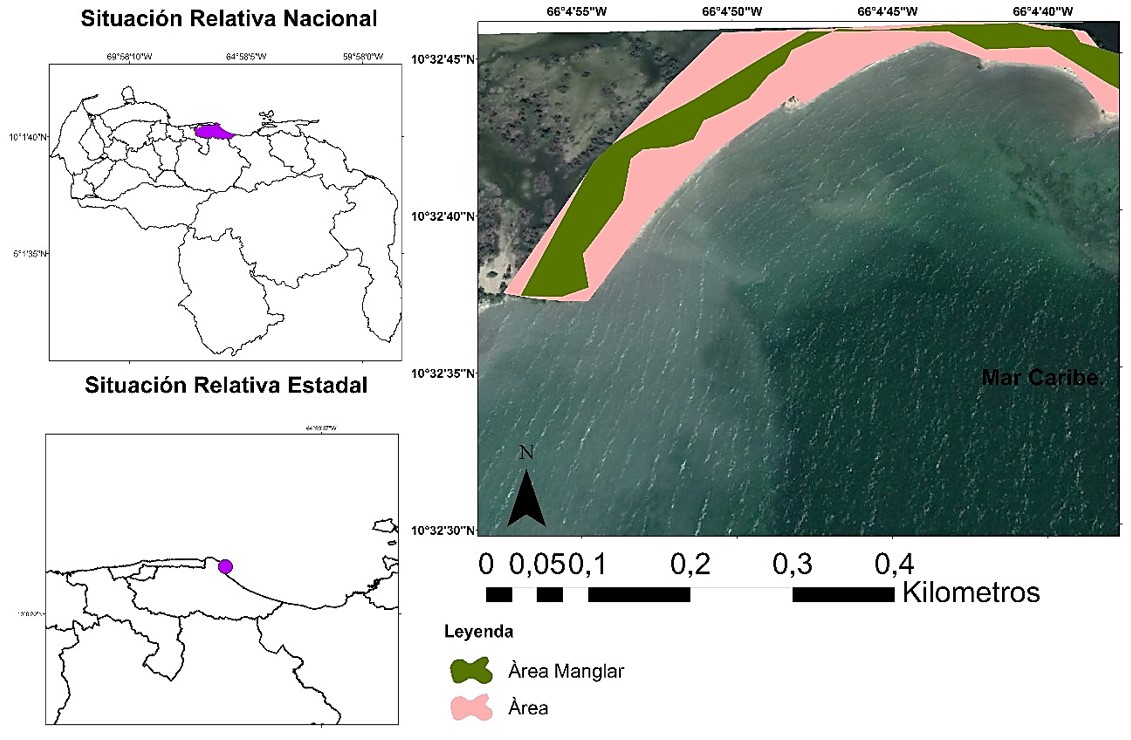
Figura 2. Mapa de la superficie cubierta por el bosque de Avicennia germinans en el sector Los Totumos, humedal Laguna Grande.
Huber (2007) y la Gobernación del Estado Bolivariano de Miranda (2010), coinciden en referir que la vegetación dominante para el tramo costero presenta espinares, palmares, herbazales y manglares. En el sector Los Totumos, se identifica el desarrollo de manglares en bosque monoespecífico de Avicennia germinans con cobertura cerrada. Lentino y Bruni (1994), reportan para el sector las desembocaduras de las quebradas: Los Totumos, Horno, Laguna Grande y Hoyo de la tierra, con un régimen tipo uadi. Adicionalmente, Lara, Suarez y Marcucci (1997), afirman que desembocan los ríos Curiepe, Capaya y Tuy, con patrones de escurrimiento permanentes.
Con base en los preceptos anteriores, se asume que el manglar ribereño y monoespecífico de Avicennia germinans emplazado sobre el suelo Entisols del sector Los Totumos en el humedal Laguna Grande, pese a que no ha sido hasta ahora objeto de estudios que permitan conocer sus condiciones estructurales, así como su aporte al ciclo total del carbono, es un espacio propicio por su accesibilidad y evolución, para identificar localmente el posible potencial forestal de la referida especie de manglar en el flujo de CO2. Es por ello que el propósito de la presente investigación fue precisar diferencias significativas en el flujo de dióxido de carbono (CO2) en suelo colonizado por Avicennia germinans así como no rizosférico, a partir del contraste de muestras superficiales (0-20 cm profundidad).
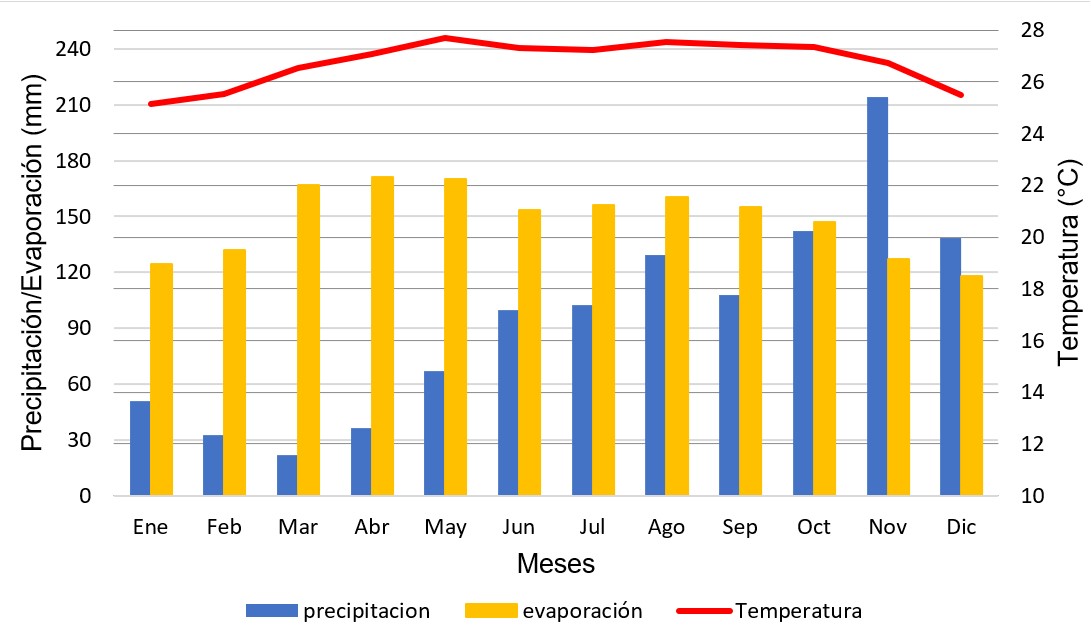
Figura 3. Climodiagrama. Elaborado a partir de la información de la hoja de datos de las respectivas estaciones, obtenidas a través de la dirección de Hidrología y Meteorología del Ministerio del Poder Popular para el Ambiente. Los datos de precipitación corresponden a la estación Carenero, y los de temperatura y evaporación a la estación Tacarigua Mamporal.
2. Materiales y Métodos
2.1 Fase de campo
El trabajo de campo se llevó a cabo en mayo de 2019, considerando que está activo el período lluvioso, lo cual constituye un aspecto fundamental en la colecta de las muestras de suelo con el propósito de cuantificar el CO2 liberado. En efecto, tal como refieren Luo y Zhou (2006), la mayor disponibilidad de agua genera un incremento en la actividad metabólica de las raíces y microorganismos, lo cual se expresa en el aumento de la respiración.
Durante la ejecución de esta fase, se realizó la observación general del área a fin de reconocer los patrones de cobertura del manglar, caracterizándolo como un bosque monoespecífico de Avicennia germinans, así como identificar la accesibilidad y homogeneidad del terreno, procurando las áreas que visiblemente presentaran el menor impacto asociado a la acción antropogénica. Con base en esta información se seleccionaron cuatro parcelas de 1000 m2, lo que representa aproximadamente el 10% del total de la superficie del bosque.
A fin de realizar la colecta de las muestras de suelo, se seleccionaron en el área del manglar dos parcelas colonizadas por Avicennia germinans, siendo estas consideradas como las que reciben la influencia de la especie como elemento interviniente en el flujo de CO2 en el suelo, y dos parcelas que aun estando ubicadas en la zona de manglar, no presentan vegetación en superficie, por lo que se consideran no rizosféricas, las cuales para efectos de la investigación constituyen la muestra sin elemento interviniente.
Las muestras de suelo se colectaron a nivel superficial (0-20 cm), considerando: (a) para zonas con presencia de Avicennia germinans, el suelo en el área más próxima a la base del individuo arbóreo, y (b) para la zona desprovista de manglar, una distancia no menor a 5 metros lineales por punto de muestreo. Bajo el criterio antes descrito, se colectaron 120 muestras de suelo, distribuidas de la siguiente manera: (a) 60 muestras de suelo dominados por Avicennia germinans, y (b) 60 muestras de suelo no rizosférico o desprovisto de manglar en superficie.
Las muestras fueron resguardadas en bolsas de polietileno denso, identificadas con etiqueta contentiva de la siguiente información: clave única de identificación, coordenadas del punto de muestreo, fecha y hora de la colecta. Las mismas fueron preservadas para su transporte al laboratorio, en una cava con una temperatura controlada entre 4 ◦C a 6◦ C.
2.2 Fase de laboratorio
Durante el desarrollo de esta fase se procedió a determinar el CO2 liberado, empleando el método de respiración basal, con base en el protocolo reportado por Anderson (1982). Para esto, las muestras de suelo conservadas a humedad de campo y cernidas en el tamiz de 10 mm fueron pre-acondicionadas a temperatura ambiente (24 ◦C), a fin de lograr su estabilización. Posteriormente, se colocaron 50 g de cada muestra de suelo en frascos de vidrio de 500 ml de capacidad con sus respectivas tapas. Adicionalmente, en frascos de vidrios color ámbar se suspendió una trampa de álcali en cada uno de los frascos, contentiva de 5 ml de una solución de Hidróxido de Sodio (NaOH) a 0,1 mol. Las muestras tratadas fueron ubicadas en un lugar oscuro del laboratorio. Transcurridas 24 horas, se procedió a retirar dicha trampa y a colocar la solución en frascos de vidrios, adicionándose con una pipeta automática 2 ml de Cloruro de Bario (BaCl2) y 2 gotas de fenolftaleína (C20H14O4) como indicador de pH para determinar la alcalinidad de la solución. El CO2 absorbido fue titulado con ácido clorhídrico (HCl) a 0,1 mol. El resultado fue expresado en mg C- CO2 g/24 h. Adicionalmente, durante la ejecución de este procedimiento también se emplearon frascos sin muestras de suelo con trampas de álcali, a fin de establecer un patrón de comparación.
2.3 Fase de análisis estadístico
Para determinar las posibles variaciones en las cuatro parcelas en estudio, así como identificar la influencia de la Avicennia germinans como fuente de variación en el flujo de CO2 en el suelo, los datos fueron analizados estadísticamente a partir de las siguientes pruebas: (a) el Análisis de varianza (ANOVA), a fin de precisar las diferencias significativas entre las medias de cada grupo de muestras, correspondientes a las cuatro parcelas en estudio; y (b) la prueba Tukey para identificar, una vez asumida las diferencias significativas en al menos uno de los cuatro conjunto de muestras, cuáles grupos de muestras son diferentes honestamente significativas.
3. Resultados y Discusión
En la Tabla 1 se presentan los valores mínimo, máximo y promedio, correspondientes al flujo de CO2 (mg C-CO2 g/24h) en el suelo para las cuatro parcelas en estudio. El ANOVA estimado por parcela arrojó para la prueba F 334,41 y una probabilidad de 6,5 × 10−57 (Tabla 2), lo que permite asumir en al menos un grupo de datos, de las cuatro parcelas en estudio, que la media es diferente con 95% confiabilidad.
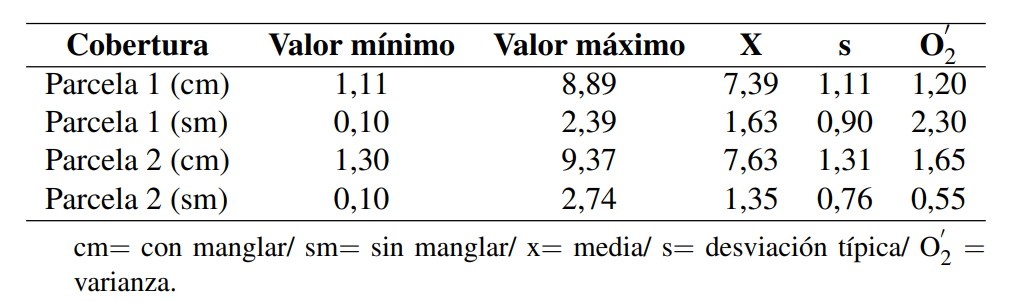
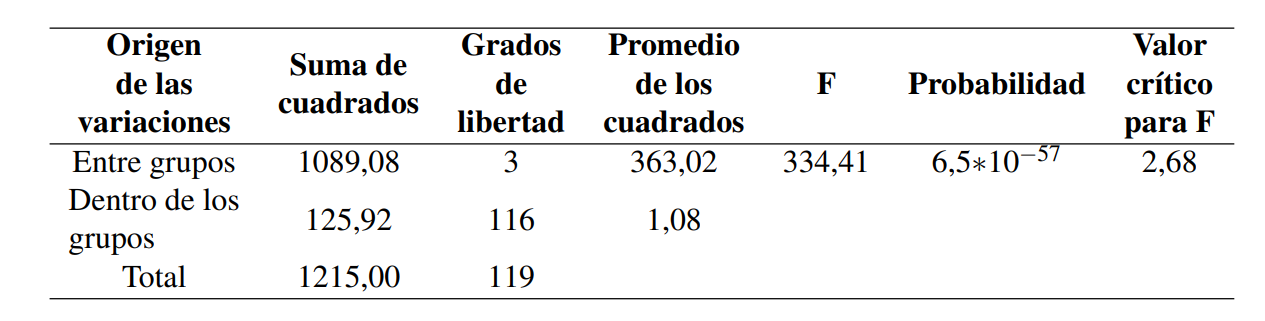
La prueba Tukey arrojó un valor de HSD de 0,69 lo que permite afirmar que: (a) no existen diferencias significativas entre las medias ponderadas de las muestras correspondientes a las parcelas con Avicennia germinans, como tampoco se identifican diferencias significativas entre las medias ponderadas de las muestras correspondientes al suelo no rizosférico, y (b) existe diferencia significativa al comparar las medias ponderadas de las parcelas que presentan manglar en superficie, con aquellas que no lo presentan (Tabla 3; Figura 4). Se considera que existe una diferencia significativa (HSD) si la diferencia de medias entre los grupos de muestras es superior al valor de HSD.

Tabla 3. Prueba de diferencia honestamente significativa de Tukey para el flujo de CO2 correspondientes a muestras de suelo en las cuatro parcelas en estudio.
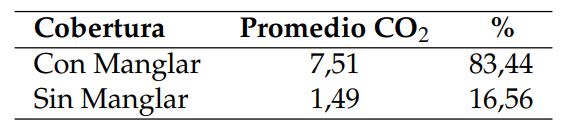
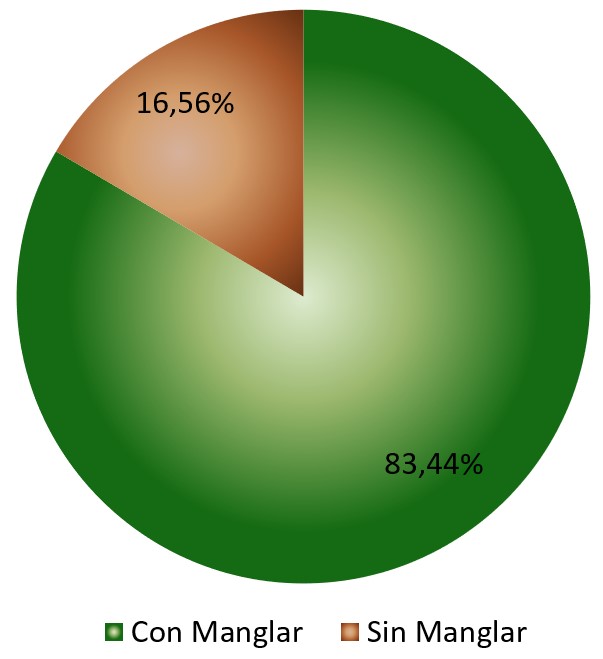
De acuerdo al análisis anteriormente referido, se puede afirmar que en la zona de estudio, el suelo cubierto en superficie por Avicennia germinans presenta un flujo de CO2 promedio de 7,51 mg CCO2 g/24h, lo cual es significativamente distinto al 1,49 mg C-CO2 g/24h estimado para los suelos desprovistos en superficie de la referida especie de manglar o no rizosféricos. Esto supone un flujo de 83,44% para el suelo cubierto por manglar, en contraste con el 16,56% estimado para el suelo no rizosférico. (Tabla 4; Figura 5). Como ya se refirió, para el sector de Boca de Uchire, Núñez y col. (2019), identificaron diferencias significativas, estimando que para el suelo no rizosférico el flujo es de 3,74 mg CCO2 g/24h, mientras que los colonizados por Avicennia germinans registran 10,61 mg C-CO2 g/24h, y en Conocarpus erectus 13,88 mg C-CO2 g/24h.
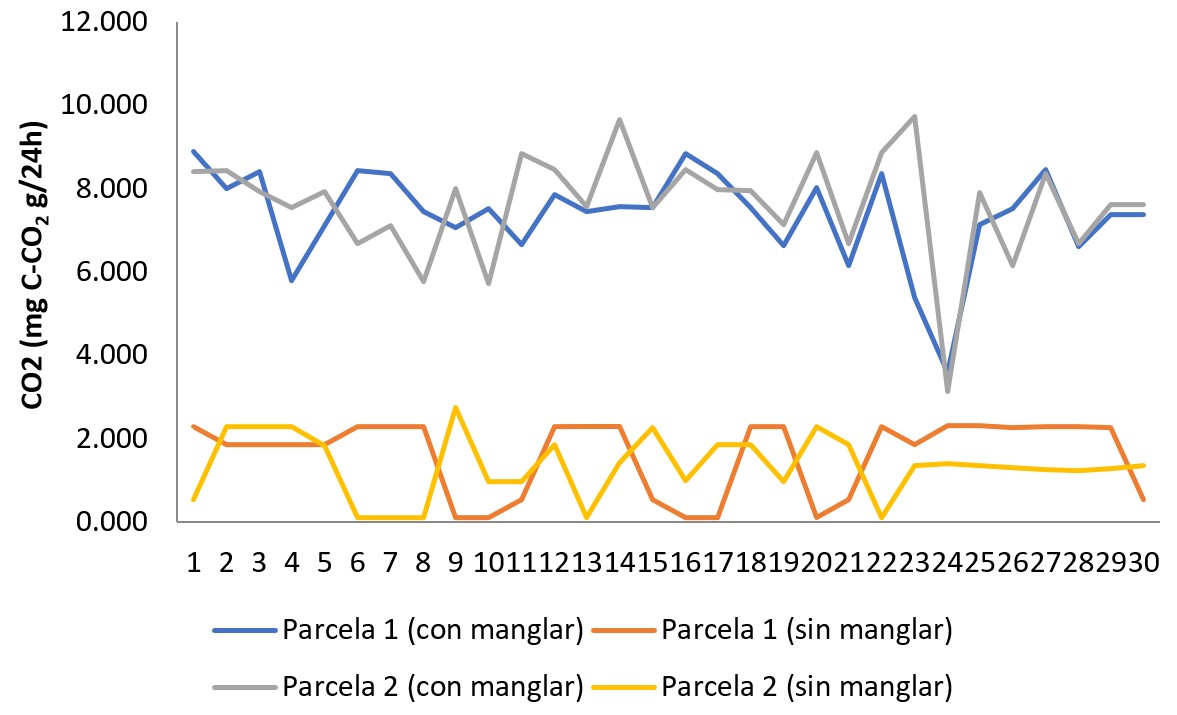
Es posible que el valor inferior de flujo de CO2 en el suelo colonizado por Avicennia germinans del sector Los Totumos, en comparación con lo reportado para la misma especie de manglar en Boca de Uchire, esté vinculado con las condiciones de salinidad edáfica, ya que el exceso de este componente no solo afecta el desarrollo estructural de las plantas (Cintron y col., 1978), limitando su potencialidad forestal como sumidero de CO2 (Mitra, Banerjee y Bhattacharyya, 2004).
En efecto, Sánchez, Paolini y Rodríguez (2010), identificaron variaciones en el flujo de CO2 en suelos colonizados por Rizhophora mangle, asociados a las fluctuaciones de sodio por períodos climáticos, registrando para el suelo altamente salino: 21,1 ± 18,9 mg C-CO2 g/24h (período seco), 25,0 ± 12,3 mg C-CO2 g/24h (período lluvioso) y 31,9 ± 18,5 (período de transición). Si bien se trata de diferentes especies de manglar, es posible que la condición de alta salinidad referida para el suelo de Los Totumos sea un factor incidente en el flujo del CO2.
Singh y col. (2010), refieren que las tasas lentas de respiración de suelo pueden ser consideradas como indicadores de una limitada actividad biológica, estrés generado por un uso inadecuado, perturbaciones climáticas, o incluso déficit de recursos. De acuerdo con Carrero y García (2009), estos manglares han sido talados para el desarrollo de obras de ingeniería con fines turísticos y comerciales, generando algunos efectos negativos. De allí que se considere que la intervención antropogénica en la zona ha generado condiciones de perturbación al equilibrio ecológico del manglar, afectando el desarrollo estructural del bosque, y limitando la potencialidad forestal en la captura y almacenamiento del CO2 en el suelo.
En cuanto a la penetración del oxígeno en el suelo en zonas de manglar, Olguín, Hernández y Sánchez (2007), refieren que éste está limitado a los primeros centímetros del perfil de suelo, debido a que el nivel freático próximo a la superficie potencia las condiciones de reducción, generando que la respiración bacteriana utilice NO3, MnO2, FeOH2, SO4 y CO2 en la zona anaeróbica como aceptores finales de electrones. De allí que el aporte de CO2 a las zonas anaeróbicas del suelo, posiblemente originado por la respiración de las raíces de los manglares, genere condiciones para la existencia de bacterias anaeróbicas, responsables de la descomposición de la materia orgánica en metano, confiriendo a esta zona del perfil del suelo la condición de sumidero de estos gases invernadero.
En la zona de oxigenación, el aporte de CO2 está posiblemente originado por la respiración de microorganismos aeróbicos que habitan en la rizosfera, constituyendo un indicador indirecto de la actividad microbiana. En efecto, Karmarkar (1982) refiere que tal actividad biológica vinculada a la proliferación de microorganismos favorece la acumulación y descomposición de residuos orgánicos aportados por el manglar, en el período en que el suelo permanece con bajo contenido de agua.
No obstante, aun en períodos de inundación, Mitsch y Gosselink (2000), afirman que tales flujos de agua ayudan a disminuir las condiciones de reducción en la zona superficial del suelo, debido a que estas láminas aportan oxígeno disuelto, lo que favorece la actividad microbiana y la consecuente liberación de CO2. Este CO2 liberado es generalmente transformado por las bacterias fotosintéticas en carbohidratos aprovechables por las plantas. Adicionalmente, los microorganismos que lo generan son fundamentales para la descomposición de la materia orgánica en humus lábil, beneficioso para el desarrollo de las plantas por su aporte de macronutrientes como el nitrógeno, indispensable para la suculencia y verdor de las hojas.
En efecto, Hesse (1961) y González y col. (2016), plantean que la Avicennia germinans requiere para su desarrollo suelos con al menos 0,4% de nitrógeno. En este sentido, Holguín y col. (2007), afirman que la proliferación de microorganismos propicia el incremento de la tasa de fijación biológica de nitrógeno, lo que contribuye con un 40 a 60% de los requerimientos del propio ecosistema de manglar.
El desarrollo de estos bosques genera un balance positivo para el sistema climático, ya que como plantean Canadell, Pitelka e Ingram (1995), presentan una alta potencialidad en el secuestro del carbono en el suelo, en respuesta al incremento del CO2 atmosférico. De allí que Zhong y Qiguo (2001), afirmaron que estos suelos deben ser considerados como reguladores naturales de la concentración y flujo de CO2 atmosférico. En efecto, Bouillon2008, afirmaron que estos bosques capturan de manera directa 4996 g CO2 m−2 por año.
Con base en los anteriores preceptos, se reconoce que la dominancia en superficie de especies de manglar contribuye con el incremento de CO2 en el suelo, y adicionalmente, se infiere que tal variación posiblemente tenga como uno de los elementos condicionantes las potencialidades edáficas de la especie colonizante, aunada a variables climáticas, edáficas y antropogénicas.
Adicionalmente, en suelos Entisols como el que se desarrolla en el sector Los Totumos, la actividad microbiana asociada a la presencia de la Avicennia germinans en superficie puede ser interpretada como un indicador del incipiente desarrollo pedogenético del mismo, ya que las bacterias activan el proceso de humificación, lo que supone que a estos suelos naturalmente minerales se adiciona por transformación de la materia orgánica, compuestos orgánicos que progresivamente aumentan la aireación, la capacidad de intercambio catiónico y las mejoras en la estructura.
4. Conclusiones
Con base en los resultados evidenciados en la presente investigación, se concluye que en el área de estudio se desarrolla un bosque monoespecífico de Avicennia germinans que cubre aproximadamente el 56% del total de la superficie del sector Los Totumos.
De igual forma, la cobertura de esta especie de manglar contribuye con el desarrollo de la actividad microbiana en la zona superficial del suelo de orden Entisols, lo cual se deduce a partir de las diferencias significativas estimadas en la respiración de muestras correspondientes a zonas rizosféricas y no rizosféricas. En efecto, para la zona colonizada por la referida especie de manglar se estimó 7,51 mg C-CO2 g/24h, mientras que la zona no rizosférica y desprovista de vegetación en superficie, la respiración estimada fue de 1,49 mg C-CO2 g/24h. Esto supone un incremento de poco más de 500% en el flujo de CO2, asociado a la presencia del manglar en la zona superficial del suelo. También, la actividad microbiana en el suelo inducida por las condiciones edáficas que genera la presencia de la Avicennia germinans posiblemente está contribuyendo con la evolución del mismo. Finalmente, se reafirma que los ecosistemas de manglar constituyen una fuente de generación de CO2, por lo que su estudio debe contribuir a generar información que permita la toma de decisiones sobre su gestión, a los fines de mitigar la emisión de este gas invernadero hacia la atmósfera.
Se reconoce que es necesario continuar los estudios en el referido manglar, a fin de profundizar en el análisis de las posibles relaciones entre sus condiciones estructurales, las propiedades edáficas del suelo en donde se emplaza y el flujo del carbono total.

