











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Forma sugerida de citar:
Jiménez Ruíz, E. R., Fonseca González, W. y Pazmiño Pesantez, L. (2019). Sistemas silvopastoriles y cambio climático: Estimación y predicción de biomasa arbórea. La Granja: Revista de Ciencias de la Vida. Vol. 29(1):45-55. http://doi.org/10.17163/lgr.n29.2019.04.
1 Introducción
El cambio climático de origen antrópico ha sido un tema muy discutido en foros a nivel mundial y los estudios que hacen referencia a este problema son abundantes (IPCC, 2013; Herrán, 2012). Las categorías de emisión más contaminantes son energía, procesos industriales y usos de subproductos (MINAE-IMN, 2014), sin dejar de lado el sector agropecuario, se conoce que la deforestación y la degradación de bosques tropicales genera entre el 15 y 35 % de las emisiones globales (Houghton, 2005). El sector agropecuario es altamente emisor por el consumo de fertilizantes, por la descomposición anaeróbica y la liberación de metano (CH 4) como el caso del arroz (MINAE-IMN, 2014) o la liberación de CH 4 generada por la ganadería (Agarwal y col., 2008). El metano es un gas de efecto invernadero con alto valor de forzamiento radiactivo 23 veces mayor al del CO 2 (Agarwal y col., 2008). Las praderas, asociadas a la ganadería ocupan el 41% de la superficie terrestre y contribuyen en un 18 % al cambio climático (9% de las emisiones de CO 2, 37% de las emisiones de metano y 65% de óxido nitroso (Steinfeld y col., 2006).
A nivel mundial se estima que existen unos 3442 millones de hectáreas con pastos (FAO, 2007), de ellas 516 millones pueden estar cubiertas de sistemas agroforestales (Nair, Mohan y Nair, 2009). La ganadería vacuna es uno de los principales usos de la tierra en América Latina, y en general, en las regiones tropicales se caracteriza por bajos niveles de productividad y rentabilidad. Y es responsable de incrementos significativos en las tasas de deforestación, acompañados de procesos de degradación de suelos, fragmentación de paisajes, pérdidas de biodiversidad, escasez del agua y reducción del nivel de ingresos (Quero Carrillo, Enríquez Quiroz y Miranda Jiménez, 2007). El sector agropecuario y especialmente los sistemas silvopastoriles son reconocidos a nivel mundial por sus múltiples beneficios, como por sus productos de carne y leche para el consumo humano, los productos de madera (madera para aserrío y leña), sus árboles que proporcionan abrigo y alimento para la fauna silvestre (Alonso, 2011); ya que contribuyen a revertir los procesos de degradación de los pastizales porque protegen el suelo, contribuyen en la recuperación de la fertilidad (Nair, Mohan y Nair, 2009), son fijadores de dióxido de carbono (CO 2) en la biomasa (Ibrahim y col., 2007; Bacab y col., 2013) y mejoran la calidad y cantidad del recurso hídrico (Nair, Mohan y Nair, 2009).
Desde el punto de vista del cambio climático son una amenaza por la liberación del metano producto de las excretas de los animales domésticos (Steinfeld y col., 2006; Agarwal y col., 2008). En América Central y alrededor del mundo, existen extensas áreas de pasturas con un alto potencial para contribuir a la mitigación del cambio climático si son manejadas bajo sistemas silvopastoriles (Nair, Mohan y Nair, 2009; FAO, 2010). Actualmente existe información científica que reconoce el potencial de mitigación de los sistemas agrícolas y ganaderos (FAO, 2010) y el tema se discute bajo la Convención Marco de las Naciones Unidas sobre Cambio Climático (Murphy y Boyle, 2012).
En las zonas semi secas de la provincia del Guayas en Ecuador sobresalen los sistemas silvopastoriles de Hyparrhenia rufa con Guazuma ulmifolia y Samanea saman. A G. ulmifolia se le atribuyen gran variedad de usos: producción de forraje y frutos para ganado bovino, frutos para fauna silvestre, tiene un néctar valioso para la producción de miel de alta calidad, leña de buena calidad, el fruto, hoja, corteza, raíz y la flor poseen propiedades medicinales, tienen múltiples efectos restauradores del medio ambiente y presta muchos servicios ambientales (Villa Herrera y col., 2009).
Con estos antecedentes, se inició un estudio cuyo objetivo fue cuantificar la capacidad de mitigación del cambio climático de sistemas silvopastoriles con G. ulmifolia, evaluando la cantidad de biomasa arbórea y bajo el suelo para desarrollar modelos de biomasa. Adicionalmente, se generó información sobre factores de expansión de biomasa e incrementos en biomasa que son elementos básicos para el monitoreo del carbono.
2 Materiales y métodos
2.1 Zona de estudio
El trabajo de campo se desarrolló en la Hacienda El Diamante, ubicada en el cantón Balzar, provincia Guayas, Ecuador, a 52 msnm, con 26°C de media anual y una precipitación media de 834, 7 a 1183, 7mm (INAMHI, 2014). El predio registra un área de 265 hectáreas, destinadas en su gran mayoría a la ganadería. La cobertura del suelo está compuesta por la combinación de samán (Samanea saman) + pasto puntero (Hyparrhenia rufa) y guamo (Guazuma ulmifolia) + (H. rufa). Guazuma ulmifolia forma parte del sistema silvopastoril desde el año 2000, cuando el propietario de la hacienda decidió proteger la regeneración de esta especie para ser usada como forraje. El área con samán, también es el resultado de una regeneración natural, luego de un incendio forestal en 1996. En ambas especies el distanciamiento entre individuos es irregular.
2.2 Evaluación de la biomasa de G. ulmifolia y de Hyparrhenia rufa
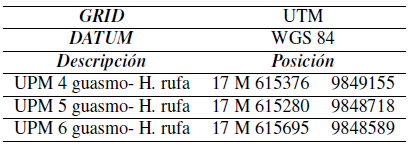
En cada uno de los sitios se instalaron tres unidades de muestreo circulares de 1000 m2 (17,84 m de radio), de forma aleatoria y se midió el diámetro a 1, 3 m sobre el nivel del suelo (Diámetro a la Altura de Pecho -DAP-) de cada uno de los árboles y la altura del fuste comercial y total con un hipsómetro Haga (50 árboles en promedio). La parcela se dividió en cuadrantes para muestrear distintos tamaños de árboles: en la unidad completa se midieron los que tenían un DAP > 4, 8cm, en 250 m 2 los arbustos entre 0;6 y 1;5 cm. El pasto puntero y la necromasa se evaluaron en unidades de 0;5 m 2. La ubicación de cada unidad de muestreo se presenta en la Tabla 1. Para calcular el volumen se usó la fórmulade Smalian (Prodan y col., 1997).
Con el fin de calcular la biomasa de hojas, ramas y raíces, se extrajo el árbol promedio por parcela (MacDicken, 1997) y se determinó en el campo el peso (kg) de cada componente. Las raíces, principalmente la de anclaje, se extrajeron con una retroexcavadora. Seguidamente se colectaron muestras húmedas de aproximadamente 1,0kg. Para el tronco se tomaron tres muestras (de la parte basal, central y apical) y se mezclaron para obtener una compuesta. Este material fue llevado al laboratorio y secado en un horno a 75°C hasta obtener un peso constante. La biomasa seca de cada componente se obtuvo del cociente “peso seco/peso húmedo”.
Con la biomasa seca de cada componente se calculó el factor de expansión de biomasa para hojas, ramas y raíz. Con el fin de aumentar el tamaño de la muestra para este cálculo se utilizó información de la misma especie, generada bajo la misma metodología, por el proyecto “Manejo del cambio climático a través del sector forestal en Costa Rica” desarrollado por la Universidad Nacional de Costa Rica. Este factor relaciona la biomasa de cada componen- te con respecto a la del fuste y fue usado para calcular la biomasa total de los árboles medidos en pie. La biomasa seca se pasó a carbono usando el factor de conversión de 0;5 (IPCC, 2013).
2.3 Ajuste de modelos alométricos
Los modelos se ajustaron mediante el método de mínimos cuadrados ordinarios con el programa estadístico Statgraphics Centurion XV. Se ensayaron aproximadamente 20 modelos para estimar la biomasa de cada componente del árbol (hojas, ramas, fuste, raíz), para el individuo completo y para el volumen. El rango diamétrico usado comprendió individuos desde 3,8 a 30,2 cm.
En todos los modelos la variable dependiente fue la biomasa o el volumen y la independiente el diámetro. En la selección de la ecuación de mejor ajuste se procedió según la metodología expuesta por Salas, (2002) y por Segura y Andrade Castañeda, (2008). Los supuestos de normalidad y homocedasticidad se probaron por medio del análisis gráfico de los residuales del modelo. Adicionalmente se graficaron los valores estimados contra los observados para ver si los modelos sobreestiman o subestiman el cálculo de la biomasa o del volumen. También por método gráfico se observó el comportamiento biológico de los modelos. La validación de las ecuaciones se realizó según lo descrito por Moret y Ruiz, (1998) y Barrales, Peña y Reguera, (2004).
3 Resultados y Discusión
3.1 Acumulación de biomasa y carbono
La cantidad de biomasa acumulada (C fijado) en sistemas agrosilvopastoriles depende de múltiples interacciones entre los componentes árbol, pasto, suelo y animal (Shibu, 2009). En los sistemas forestales el carbono se acumula en cuatro componentes (biomasa arriba del suelo, necromasa, sistemas radiculares y carbono orgánico del suelo) y la biomasa leñosa representa el carbono con mayor permanencia (Snowdon y col., 2001) y el mayor almacén a nivel de árbol.
En el presente estudio la biomasa total fue de 19, 75 Mgha −1 (9, 8 Mg C), los árboles de guasmo según el muestreo acumularon el 83 % (16, 45 Mg ha −1; 8, 23 Mg Cha −1), mientras que el aporte de la necromasa y el pasto fue de 1, 4 y 1, 9 Mg ha −1 (0, 7 y 0, 95 Mg Cha −1), con incrementos medios de 1, 08 Mg ha −1 ao −1 y 0, 55 Mg ha −1 ao −1, respectivamente. El componente arbóreo se compone de un promedio de 497 árboles ha−1, distribuidos en una amplitud diamétrica de 3, 8 a 30, 2 cm.
Estas reservas de biomasa y/o carbono son comparables a los existentes en otras latitudes en bosques caducifolios, por ejemplo, en el trópico seco de Costa Rica, en sistemas silvopastoriles las especies maderables nativas Dalbergia retusa, Dyphisa robinioides y Pithecellobium saman en la biomasa arbórea aérea fijaron anualmente 0;2; 1;25 y 0;26 Mg Cha -1, respectivamente (Rojas, Ibrahim y Andrade, 2009). En zonas de clima seco en México se han reportado valores promedios de biomasa de árboles en potrero de 13 Mg ha −1 (Hughes Flint, Kauffman Boone y Jaramillo, 2000) y de 5, 9 a 7, 7 Mg ha −1 («Biomass, carbon, and nitrogen pools in Mexican tropical dry forest landscapes»). Torres Rivera y col., (2011) reportan para un sistema silvopastoril con baja densidad del componente arbóreo (120 árboles ha −1) una acumulación de 2, 86 Mg Cha −1 a los tres años. En Colombia, con la implementación de sistemas silvopastoriles Chará y col., (2009) citan incrementos en C almacenado de 0, 51 Mg ha −1 ao −1 y en Costa Rica a los cuatro años y tres meses 12, 5 Mg Cha −1 (Andrade, Brook e Ibrahim, 2008). También en potreros arbolados sin una edad definida (Chacón León y Alice Harvey, 2013) para Costa Rica (Esparza) y Nicaragua (Mitiguás) indican un promedio de biomasa de 10;7 Mg ha -1 con variaciones entre 5;3 y13;5 Mg ha -1, para estos mismos sitios Ibrahim y col., (2007) cita cifras de 3;2 a 14;2 Mg ha -1 en árboles dispersos en potreros en Esparza y de 9 a 17;9 Mg ha -1 en Matiguás.
En Nicaragua, Ruiz, (2002) encontró contenidos de carbono de 8, 2 3, 0 Mg Cha −1 en pasturas nativas con árboles (100 árboles ha −1) y en pasturas mejoradas con árboles (110 árboles ha −1) de 12, 5 3, 6 Mg ha −1. Mientras que en los Andes Colombianos la biomasa aérea de Acacia decurrens en sistema silvopastoril con abundancias de 1111 y 407 árboles ha −1, mostró un incremento un anual de 9, 9 y 11, 2 Mg Cha −1 (Giraldo, Zapata y Montoya, 2008).
En el componente arbóreo la raíz acumuló el 18% de la biomasa total (2, 96 Mg ha−1) y los componentes leñosos -raíz, fuste y ramas- el 95% (15, 63 Mg ha −1), similar a lo citado por Anguiano, Aguirre y Palma, (2013) quienes reportan cifras entre 85 y 98% incluyendo las hojas. La biomasa aérea de los árboles fue de 13;49 Mg ha-1. El tronco representa el 47;4% de la biomasa del árbol y el 57;7% de la biomasa aérea, valor que concuerda con lo encontrado por Gómez y col., (2010) para Gliricidia sepium (60;9%) y Leucaena leucpcehala (58;3%).
La biomasa (C) determinada en este estudio también es similar a la reportada en sistemas agroforestales en Changuinola Panamá, en cacao plantado a 33 m más laurel a 66 m y 1212 m; el Mg Cha −1, cuyo incremento anual fue de 1, 7 y 2, 5 Mg Cha −1 (Ortiz, Riascos y Somarriba, 2008). Según Umaña y Conde, (2013), en Tolima, Colombia, en sistemas agroforestales de aguacate + plátano, cacao + aguacate y cacao + plátano, la fijación anual de carbono fue 2, 23; 4, 14 y 0, 52 Mg, respectivamente
En el componente arbóreo, la raíz a pesar de representar un carbono de mayor permanencia, ha sido poca estudiada porque se le atribuye un alto grado de dificultad (Dixon, 1995; Schlegel, 2001), a pesar de representar entre el 10 y 40 % de la biomasa total (Cairns y col., 1997; MacDicken, 1997; Andrade e Ibrahim, 2003), en este estudio la raíz acumula el 18 % de la biomasa del árbol. Cuantificar la biomasa radical es un esfuerzo que debe reconocerse en este estudio por la poca disponibilidad de información existente para la diversidad de especies. Usar valores generales en vez de información local y específica por especie puede generar cálculos con mucha imprecisión.
Debe tenerse presente que, aunque la cantidad de biomasa o carbono determinada en este estudio sea similar o esté dentro del rango de lo reportado en la literatura, siempre existirán diferencias influenciadas por la diversidad de arreglos espaciales de los sistemas silvopastoriles, diversidad de especies y variaciones metodológicas para cuantificar la biomasa.
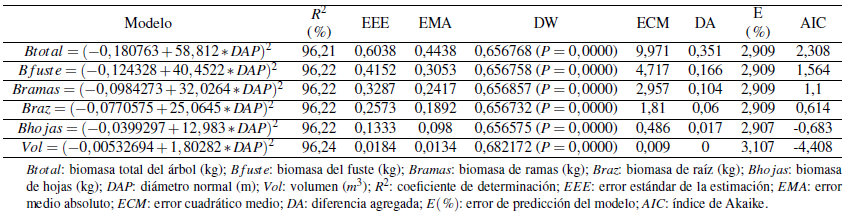
3.2 Modelos de biomasa, volumen y factores de expansión
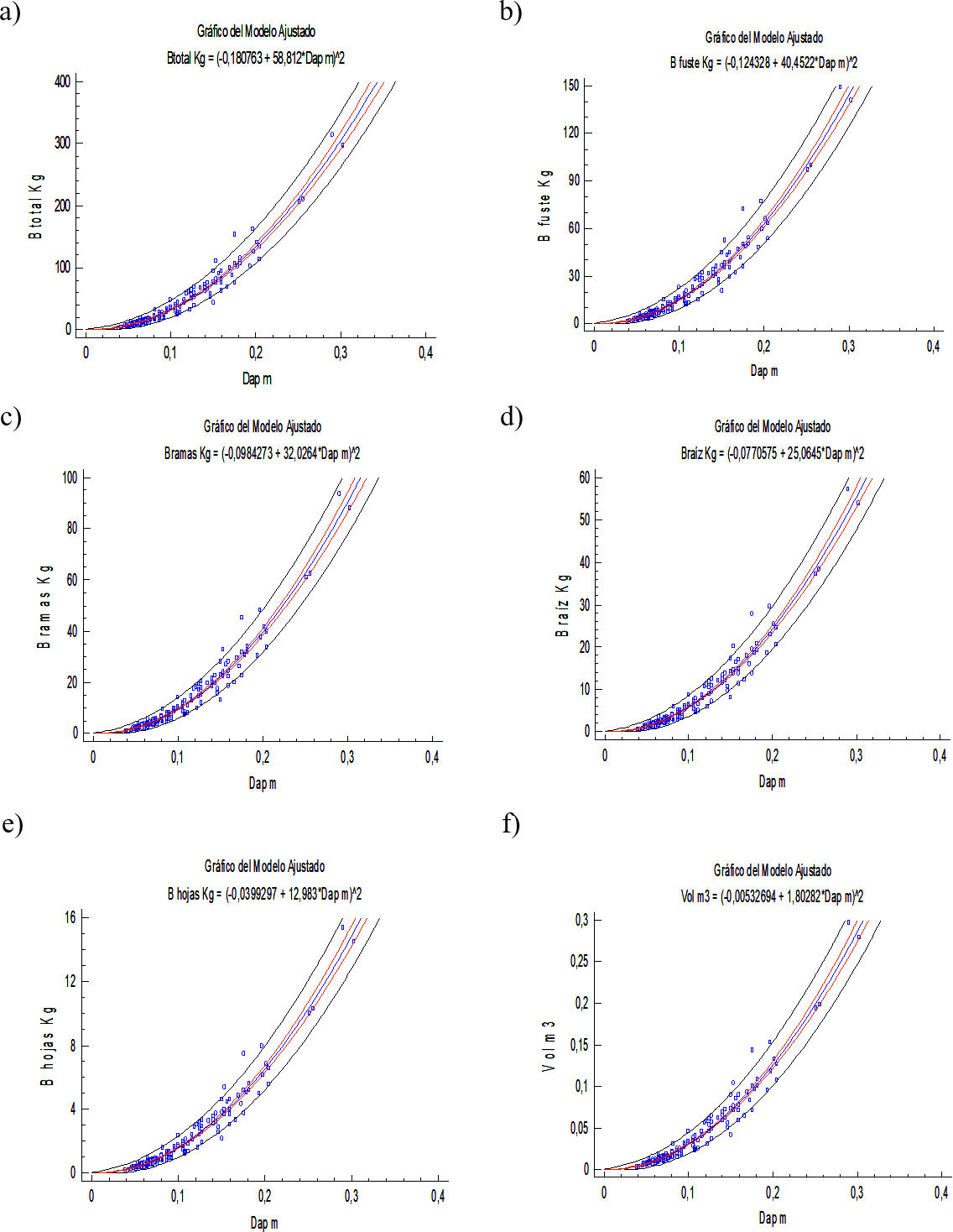
Los modelos para estimar la biomasa de los distintos componentes del árbol y para la biomasa total presentaron muy buenos ajustes (R2 > 0, 96, con P < 0, 0001) y valores muy bajos para los otros estadísticos. Es importante resaltar que el error de predicción no supera el 3,1 % (Tabla 2, Figura 1 y 2). Además, son modelos muy sencillos y prácticos que presentan poco grado de dificultad para el cálculo de la biomasa por usar solo el diámetro como variable regresiva. Se han logrado características similares de ajuste (predicción) con modelos para árboles en sistemas silvopastoriles cuya arquitectura es diferente a la de árboles en plantación pura y de bosque natural, por ejemplo, para árboles dispersos de motilón silvestre Freziera canescens Cabrera y col., (2007) logró una ajuste de R2 = 89, 61 para estimar la biomasa aérea en función del DAP, y ajuste más bajo 71, 81; 87, 60; 87, 57, cuando usaron como variable independiente la altura, el volumen y el área basal, respectivamente. Gómez y col., (2010) lograron ajustes inferiores lograron para Gliricidia sepium (R2 entre 75 y 86) y de 61 a 87 % en Leucaena leucocephala.
Es sorprendente el buen ajuste para ramas y hojas, componentes que en otras investigaciones presentan un ajuste inferior como el reportado por Fonseca, Alice y Rey, (2009) para especies en bosque secundario (83,04 % en hojas, 88, 25 % en raíz), para Vochysia guatemalenis (89,05 % en hojas) e Hieronyma alchorneoides (89, 69 % en raíz), o R2 superiores al 70%, con errores de estimación superiores al 25% (Fonseca y col., 2016). Correlaciones bajas entre la biomasa de hojas y ramas con el diámetro también fueron reportadas por Montero y Kanninen, (2002), Návar, González y Graciano, (2011) y Pérez y Kanninen, (2003).
El modelo elegido para estimar el volumen total del fuste presenta las mismas características de los modelos para biomasa (Tabla 2, Figura 1 y 2). La bondad de ajuste lograda (R2= 96, 2 %) es igual o superior al obtenido para otras especies (Da Cunha y Guimarães Finger, 2009; Barrios, López y Nieto, 2014; Ramos y col., 2014).
El factor de expansión de biomasa para hojas fue de 0,1 (S = 0, 03), 0,66 (S = 0, 15) para ramas y de 0, 39 (S = 0, 10) en raíz, valores que se encuentran dentro del rango reportado en diferentes estudios, con diferentes especies y para otras latitudes (Fonseca, Alice y Rey, 2009; Schlegel, 2001; Segura y col., 2000).
4 Conclusiones
Los sistemas silvopastoriles representan un aporte significativo a la mitigación del cambio climático, beneficio que puede incrementarse si se introducen técnicas de manejo.
Las ecuaciones para estimar la biomasa y el volumen de los árboles ayudan a reducir el costo de los inventarios de carbono para aquellas organizaciones que están en proceso de buscar la neutralidad del carbono o para la definición de políticas a nivel nacional. El uso del diámetro como variable predictora el diámetro facilita su aplicación y reduce los errores de estimación, además, la capacidad de predicciónde estos modelos (R2 > 96%) y el bajo errorde estimación (E < 3%) aportan mucha confiabilidaden las estimaciones.
Cuantificar la biomasa radical es un esfuerzo quedebe reconocerse en este estudio por la poca disponibilidad de información existente para la diversidad de especies. El uso de valores generales en vez de información local y específica por especie puede generar cálculos con muncha imprecisión.
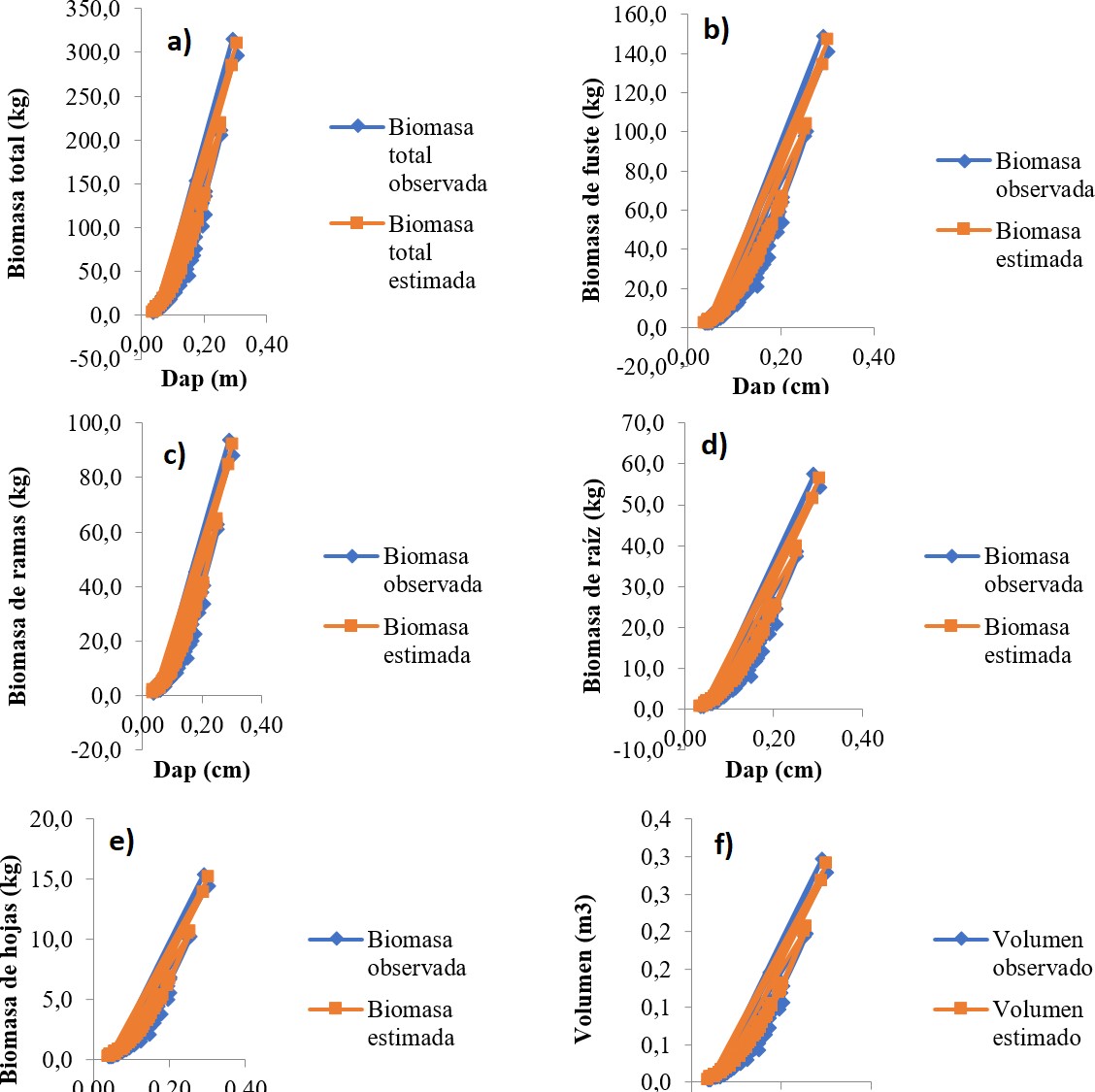
Figura 2. Estimación de biomasa y volumen a partir del diámetro: a) biomasa total [Btotal = (−0,180763 + 58,812 ∗ DAP)2], b) biomasa del fuste [B f uste = (−0,124328 + 40,4522 ∗ DAP)2], c) biomasa de ramas [Bramas = (−0,0984273 + 32,0264 ∗ DAP)2], d) biomasa de raíz [Braz = (−0,0770575 + 25,0645 ∗ DAP)2], e) biomasa de hojas [Bhojas = (−0,0399297 + 12,983 ∗ DAP)2], f) volumen del fuste [Vol = (−0,00532694 + 1,80282 ∗ DAP)2].

