











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El bosque seco neotropical destaca por su extensión, biodiversidad y servicios ecosistémicos (Calvo-Rodriguez y col., 2017; Hubbell, 1979). Estos bosques han sufrido un alto impacto antropogénico en las últimas décadas (Aguirre y col., 2017). Particularmente en Ecuador, país del neotrópico con más especies vegetales catalogadas al menos como vulnerables (IUCN, ), se estima que al menos el 50 % de la extensión de bosque seco ha desaparecido en las últimas décadas por la intervención humana (Aguirre, Kvist y Sánchez, ), principalmente debido a la deforestación y transformación del suelo a terreno agrícola y ganadero (Madriñán, 2014; Aguirre y col., ; Aguirre y Kvist, ). El impacto al que están sujetos estos bosques contrasta con que son de los ecosistemas menos estudiados en las diversas escalas ecológicas (Aguirre y col., ). Para identificar, establecer y mantener acciones para la restauración y conservación es por tanto necesario ampliar el conocimiento biológico de al menos las especies vegetales más relevantes (Aguirre, Kvist y Sánchez, ).
Una de las especies más relevantes del bosque seco de Ecuador es Cavanillesia platanifolia (Humb. y Bonpl.) Kunth (Aguirre y col., 2017; Espinosa y col., 2011), especie conocida comúnmente en América Central y del Sur como mocundo de Cartagena, bongo/a, macondo, cuipo, güipo, hameli, hamelí, pretino, pigio o pijío. Su distribución se ciñe principalmente a los bosques secos tropicales del corredor Chocó-Darién, desde Panamá hasta el norte de Perú, aunque está también presente en bosques húmedos.
Este árbol de la familia de las Malváceas-Bombacáceas puede superar los 40 metros de altura (Figura 1A), y destaca en Ecuador su rol como generador de hábitat, entre otros para la anidación del papagayo de Guayaquil Ara ambigua guayaquilensis (López-Lanús y Socola, 2000), y el aporte nutricional de su semilla, la cual es comestible y recurso de la fauna local (Adler, 1995; Madriñán, 2014). También cabe resaltar su trascendencia a nivel paisajístico y estructural (Aguirre, 2012). Es usada como recurso maderero por las comunidades locales en diversos países como Panamá (Correa, Galdames y De Stapf, 2004), pero no hay constancia de ello en Ecuador (Madriñán, ). La especie ha sido declarada como amenazada globalmente en América Central (IUCN, ). En Colombia, donde ha sido indexada como una de las especies más importantes del bosque seco tropical (Ruiz y Saab, 2020), se considera en peligro (IUCN, ). Su distribución potencial en este país puede no estar severamente amenazada por el cambio climático, aunque sí se prevé que pueda diezmar sus poblaciones (Aguirre y col., ).
Muchas características son comunes entre las distintas especies del género, como la esciofilia; las plántulas (Figura 1B) y juveniles crecen en lugares donde la radiación solar es escasa (Vieira, Scariot y Holl, 2007; Montalvo, Rommel y Cevallos, 2013). Por lo general, estas especies se caracterizan por su baja densidad (Vieira y Scariot, ; Vieira y col., ; Melo-Júnior y col., 2015), rápido crecimiento (Condit, Hubbell y Foster, 1993), y su tolerancia a la sequía durante la estación seca (Wolfe, 2017). En septiembre-octubre producen sus frutos grandes y carnosos, que contienen una semilla anemócora con cáscara externa que contiene blando mucílago (Vieira y Scariot, ). Los rasgos de las semillas se han estudiado sobre todo en C. arborea. Las semillas de esta especie rondan los 8g de peso fresco, mientras que el peso seco es de alrededor de 1g (Vieira y Scariot, ; Romero-Saritama, 2016). Las semillas con el tiempo pierden viabilidad (Walters, Wheeler y Grotenhuis, 2005) y son sensibles a la desecación (Vieira y Scariot, ; Vieira, Scariot y Holl, ; Vieira y col., ; Lima y col., 2008). Como en gran parte de las especies del bosque tropical seco, la baja humedad mantiene en latencia las semillas, la cual puede interrumpirse durante la estación lluviosa que activa el embrión para su germinación (Debeaujon, Leon-Kloosterziel y Koornneef, 2000).
Cuando se han muestreado los individuos de C. platanifolia en los bosques secos de Ecuador para poner en marcha planes de conservación y restauración, el dato más relevante es la baja presencia de plántulas y juveniles, lo cual indica una muy baja tasa de regeneración natural (Villalba-Briones, observación personal). La ausencia de individuos jóvenes de C. platanifolia no solo ocurre en las proximidades de individuos adultos, lo cual muchas veces se relaciona con una alta mortalidad densodependiente debida a patógenos, depredadores de semillas y/o herbivoría que tiene lugar bajo la copa materna (Alvarez-Loayza y Terborgh, 2011; Comita y col., 2014; Xu y col., 2015), sino que es constante a lo largo y ancho de los bosques. En otras especies de Malváceas-Bombacáceas del neotrópico como Pachira quinata (Jacq.) Alverson también se ha observado bajas tasas de regeneración natural, lo cual se debe a la disminución en la producción de frutos, a una intensa depredación de frutos y a la mortalidad de plántulas debido a patógenos (Castellanos y Stevenson, 2011).
En cuanto a la depredación de semillas, se conoce que en otras especies del género Cavanillesia, como C. arborea (Willd.) Schum. y C. chicamochae Alonso, sus semillas son intensamente depredadas tanto pre-dispersivamente (insectos varios y loros de la familia Psittacidae) como post-dispersivamente (roedores y otros mamíferos), y que la depredación de semillas puede incrementarse notablemente con el silvopastoreo (Vieira y Scariot, ; Vieira, Scariot y Holl, ; Díaz-Pérez, Puerto-Hurtado y Fernández-Alonso, 2011; Souza-Silva y col., 2015). Si bien la actividad silvopastoril no es tan intensa en los bosques secos de Ecuador (Aguirre, 2012), la tasa de depredación de semillas, particularmente por roedores, puede haber aumentado por la continua pérdida de mamíferos carnívoros (Catterall, 2018) y por la defaunación de grandes herbívoros debido a la presión que la caza ejerce (Rosin y Poulsen, 2016). De manera más importante, y al igual que se ha observado para C. arborea y C. chicamochae, otras actividades antrópicas como la fragmentación del bosque y la reconversión del suelo pueden estar disminuyendo la emergencia y el reclutamiento de las semillas de C. platanifolia (Vieira y Scariot, ; Vieira, Scariot y Holl, ; Melo-Júnior y col., 2015; SouzaSilva y col., ).

Figura 1 Cavanillesia platanifolia (Humb. & Bonpl.) Kunth (A) Ejemplar del punto de muestreo en la provincia del Guayas (Ecuador), con sus característicos anillos transversales en el tronco. (B) Plántula en vivero con sus hojas simples y trilobuladas.
Conocer aspectos básicos de la biología como la viabilidad de la semilla y su capacidad de reclutamiento son en sí mismos objeto de investigación necesarios para la conservación de cualquier especie. En las especies tropicales, las semillas son componentes críticos de la reproducción, y los factores que influyen en su persistencia, supervivencia y emergencia son determinantes para la producción de plántulas y árboles jóvenes en los que se basan los planes de restauración (Vieira y Scariot, ; Sarmiento y col., 2017). Particularmente, es indispensable el éxito en la conservación de la semilla para planificar y desarrollar el cultivo de la especie en vivero como estrategia de conservación ex situ (Becerra-Vázquez y col., 2018). En el presente estudio evaluamos la posibilidad de que la conservación en frío de las semillas de C. platanifolia pudiera favorecer su emergencia, supervivencia y rendimiento.
Materiales y Métodos
Área de estudio
El bosque seco en Ecuador se encuentra en el flanco occidental de los Andes, en las provincias de Esmeraldas, Manabí, Guayas, El Oro y de Loja (Aguirre y col., 2017). Cavanillesia platanifolia suele encontrarse en estos bosques por debajo de los 800 m s.n.m. (Jørgensen y León-Yanes, 1999; Aguirre, Kvist y Sánchez, ). Según Espinosa y col. (2011), se distribuye por las provincias de El Oro, Loja, Manabí, Guayas y en el bosque seco tumbesino, así como en la provincia de Esmeraldas (Aguirre y col., ).
Para el presente estudio, se recolectaron semillas de C. platanifolia en la Hacienda Pabeclar, ubicada en el Bosque Protector Chongón (2◦ 7’ 13.97"S, 80◦ 4’ 57.78.O; provincia de Guayas, Figura 2). Esta región, limitante con la cordillera Chongón-Colonche y que es parte del corredor Chocó-Darién de la costa de Ecuador, es objeto prioritario de conservación (Mittermeier y col., 2011). En el área forestal en cuestión la actividad agrícola va en aumento, principalmente debido a la creciente extensión de los cultivos de maíz (Figura 2). La recolección de semillas se realizó en noviembre del 2017, en un área de aproximadamente 6000 m 2, seleccionando aquellas del suelo que no presentaban ningún signo de depredación (Bonfil-Sanders, Cajero-Lázaro y Evans, 2008; Orantes-García y col., 2013). Dentro del área de recogida se caracterizaron los individuos (N=6) de C. platanifolia, que en esta zona tenían una circunferencia a la altura del pecho (CAP) a 1.30 m del suelo de 321 cm a 570 cm y una altura de 25 a 37 m, medidas mediante la técnica Range Finder (Wing, Solmie y Kellogg, 2004).
Metodología de conservación, metodología de vivero, siembra de las semillas, y seguimiento de la emergencia y rendimiento de las plántulas
Ensayo piloto
Se realizó un ensayo piloto en invernadero con 187 semillas (iniciando la hidratación de la semilla el 28 de febrero del 2018), para comprobar la viabilidad de las semillas recolectadas. Previo a la siembra, desde la recolección previamente mencionada, las semillas se almacenaron en un espacio cerrado, en oscuridad y temperatura ambiente (21,3◦C- 26,8◦C) durante 3 meses. Después, las semillas se sometieron al proceso de escarificación física, y posteriormente se dejaron en remojo durante 48 horas. Según Trujillo (1995), esta hidratación no debe ser inferior a dos horas, pero tampoco superior a las 48 para evitar posibles fermentaciones. Tras la hidratación, las semillas fueron dispuestas en cuadros de 1 m 2 x 30 cm de alto con 2 cm de distancia entre ellas. Debido a la alta mortalidad de las plántulas de C. platanifolia a causa de patógenos (Villalba-Briones et al., observación personal), se utilizó un sustrato de arena con alto contenido en sílice, el cual tiene efecto fungicida. Los cuadros se dispusieron en un vivero (invernadero) con malla sarán, la cual reduce la radiación solar en un 80 % y favorece la emergencia de las semillas de otras especies del género (Vieira y col., 2008). A partir de entonces, los cuadros se regaron en el vivero 5 días a la semana durante 10 minutos por nebulización. Desde el momento de la siembra, y a lo largo de 1 mes, se midió la tasa de emergencia acumulada, la cual fue de un 89.3 %.
Ensayo de respuesta a condiciones de conservación
Con semillas del mismo origen y fecha de recolección, procedimos con el ensayo experimental, con un tiempo de almacenamiento más prolongado que en el ensayo piloto. Para este ensayo, primero las semillas recolectadas de C. platanifolia se almacenaron en seco en un espacio cerrado salvaguardado de los elementos externos ambientales por un total de 3 meses (noviembre del 2017 a febrero del 2018), tras los cuales fueron aleatoriamente asignados a 2 tratamientos con 55 semillas por tratamiento. Para el primer tratamiento, las semillas se mantuvieron almacenadas durante 6 meses más a temperatura ambiente (21,326,8◦C) y oscuridad. Para el segundo tratamiento, las semillas se almacenaron en un refrigerador (4,3-7,6 ◦C) por el mismo tiempo. Mediante el tratamiento de conservación en frío la intención era comprobar si la conservación favorece las semillas manteniendo su viabilidad. Este hecho ha sido sugerido por varios autores (Becerra-Vázquez y col., 2018; Li y Pritchard, 2009), aunque también se ha observado una alta variabilidad intraespecífica (Posada, Ocampo y Santos, 2014; Trujillo, 1995).
Esta observada variabilidad intraespecífica se debe en gran parte a las características morfológicas y fisiológicas de las semillas, como su contenido hídrico y termo-tolerancia (Kranner y col., 2010; Sasaki, 2008). Para nuestro ensayo experimental optamos por la conservación en refrigerador por ser económica y fácilmente reproducible, en vista de que la conservación en nitrógeno líquido (-196◦C) no incrementa significativamente la emergencia en otras especies como C. arborea (Lima y col., 2008). Tras los 6 meses de tratamiento y previo a la siembra, las semillas se sometieron al proceso de escarificación física, y posteriormente se dejaron en remojo durante 24 horas. Tras la hidratación, procedimos a la siembra y cultivo en vivero (invernadero) de igual manera a lo descrito para el ensayo piloto. Al mismo tiempo, la progresión de desarrollo de la semilla fue registrada fotográficamente en un recipiente de vidrio adaptado para la recolección de imágenes (Figura 3). Desde el momento de la siembra, y a lo largo de 1 mes, medimos la tasa de emergencia acumulada y mortalidad de las plántulas emergidas.
A partir de estas variables también estimamos el éxito de reclutamiento (emergencia + supervivencia). A su vez, medimos el crecimiento (altura) de las plántulas a los 14 y a los 25 días de la siembra (ver ejemplo de plántula en la Figura 4). Durante todo el proceso se midieron los parámetros ambientales de temperatura, humedad relativa y punto de rocío. Las mediciones se realizaron con un termohigrómetro, tomando 3 medidas por cuadro/día para hacer la media diaria. Las medidas fueron homogéneas a lo largo del tiempo, con una variación de 3◦C en temperatura (27,3-30,3◦C) y de 2,8◦C en punto de rocío (21,7-24,5◦C). En cuanto a la humedad relativa, no osciló más allá del 25 % entre las distintas mediciones (65,1- 85,3 %).

Figura 3 Cavanillesia platanifolia (Humb. & Bonpl.) Kunth (A) Semilla recolectada con sus 4 alerones para ser dispersada anemócoramente. (B) Semilla una vez descascarillada. (C) Semilla hidratada a las 24 horas. (D) Posicionamiento en arena de una semilla para su germinación. (E) Semilla germinando a las 48 horas. (F) Emergencia de la plántula.
Análisis estadístico
Analizamos el efecto de ambos tratamientos (conservación en frío vs. conservación a temperatura ambiente) sobre la emergencia, mortalidad y éxito de reclutamiento de cada semilla con modelos lineales generalizados con distribución binomial. En cuanto al efecto de los tratamientos sobre el crecimiento (altura) de las plántulas en ambas mediciones temporales (a los 14 y a los 25 días de la siembra), evaluamos si se cumplían los parámetros de normalidad. Los gráficos de Cuantil-Cuantil determinaron que los datos se ajustaban bien a una distribución normal, y el test de normalidad ShapiroWilk determinó que no se descartara la hipótesis nula de la asunción de normalidad en ambos casos. Por tanto, la altura de las plántulas pudo ser analizada por medio de un modelo lineal general (ANOVA de un factor). Los análisis se realizaron con el software estadístico R 3.5.2 (R Core Team, 2017), mediante los paquetes nlme (Pinheiro y col., 2011) y lme4 (Bates y col., 2014).

Resultados y Discusión
En el ensayo experimental, las semillas conservadas en frío tuvieron una tasa de emergencia 10 % mayor que las conservadas a temperatura ambiente (98,18 ± 1,81 vs. 89,09 ± 4,24 %, χ2 = 4,20, P = 0,040; Figura 5A) y, además, la tasa de mortalidad con la conservación en frio fue 317 % menor una vez emergidas (16,66 ± 5,11 vs. 71,42 ± 6,52 %, χ2 = 33,30, P <0,0001; Figura 5B). La mortalidad estuvo severa mente influenciada por infecciones bacterianas y sobre todo fúngicas que afectaron el endospermo y los cotiledones. El éxito de reclutamiento de las semillas, al considerar conjuntamente la emergencia y supervivencia, fue un 228 % mayor en las semillas conservadas en frío respecto a las conservadas a temperatura ambiente (81,81 ± 5,24 vs. 25,45 ± 5,92 %, χ2 = 37,35, P <0,0001; Figura 5C).
Además de un mayor reclutamiento, las plántulas de semillas conservadas en frío mostraron un mayor rendimiento durante el seguimiento efectuado el mes siguiente de la siembra. Así, las plántulas emergidas de las semillas conservadas en frío resultaron ser un 45 % más altas que las conservadas a temperatura ambiente a los catorce días (8,60 ± 0,42 vs. 5,92 ± 0,73 cm, F1,58 = 9,53, P = 0,003; Figura 5D), y un 28 % más altas a los veinticinco días (17,10 ± 0,55 vs. 13,29 ± 1,28 cm, F1,57 = 9,74, P = 0,002; Figura 5D).
Diversos factores están diezmando las poblaciones de Cavanillesia platanifolia en el neotrópico. A la pérdida y fragmentación del área forestal y a su tala se une la recientemente observada escasa regeneración natural en los bosques secos de Ecuador. Ello puede deberse una disminución en el número de individuos y una menor producción de frutos y semillas, a una mayor tasa de depredación de semillas, a un incremento en la desecación de las semillas y/o en la mortalidad de las plántulas debido a factores ambientales. Así, el establecimiento de los individuos jóvenes puede estar viéndose comprometido por un creciente estrés fisiológico debido a las cada vez mayores oscilaciones climáticas estacionales. Este hecho podría agravarse en el futuro, por lo que sería recomendable estudiar de manera completa el ciclo biológico de C. platanifolia hasta su etapa adulta, y determinar la influencia de los distintos factores bióticos y abióticos sobre su eficacia biológica. No puede obviarse que la viabilidad de las semillas también puede estar disminuyendo por menores tasas de polinización cruzada (Hamrick y Murawski, 1990).
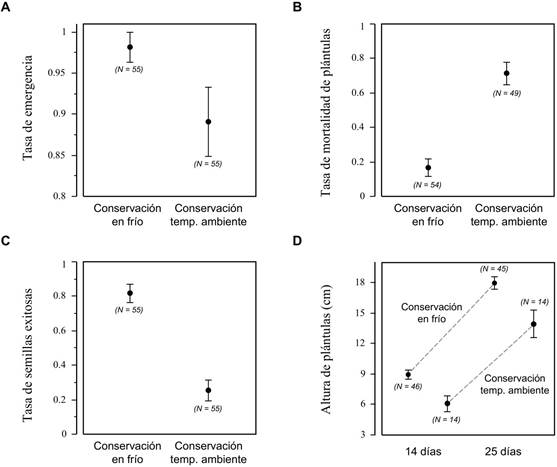
Figura 5 Efecto de la temperatura de conservación de las semillas de Cavanillesia platanifolia (Humb. & Bonpl.) Kunth. sobre las variables observadas. (A) Tasa de emergencia. (B) Tasa de mortalidad de plántulas. (C) Tasa de reclutamiento (semillas exitosas). (D) Altura de las plántulas con diferentes tratamientos a los 14 y a los 25 días de la siembra.
El presente estudio demuestra que la conservación en frío de las semillas durante seis meses permitió el mantenimiento de su viabilidad respecto a las semillas conservadas a temperatura ambiente, incrementando la tasa de emergencia. La conservación en frío permitiría por tanto mantener la viabilidad de semillas sensibles a la desecación (Normah, Sulong y Reed, 2019), como es el caso de C. platanifolia. Nuestro estudio demuestra que la conservación en frío de las semillas durante 6 meses mantuvo significativamente la viabilidad de las semillas. La emergencia de las semillas conservadas en frío durante seis meses (con tres meses previos de conservación a temperatura ambiente en un lugar cerrado) fue similar, y e incluso ligeramente superior a la emergencia observada en semillas de tres meses a temperatura ambiente en el mismo lugar cerrado (ensayo piloto), efecto posiblemente mediado por la interrupción de la dormancia debida al frío.
Además, es destacable que las plántulas provenientes del tratamiento de conservación en frío crecieron de manera más vigorosa. Este tipo de conservación limitaría los efectos adversos de las altas temperaturas y del déficit de agua sobre las semillas, disminuyendo su envejecimiento. Mediante el fenómeno multicausal del envejecimiento se reduce la calidad fisiológica de las semillas induciendo cambios estructurales, composicionales y funcionales, limitando su viabilidad y vigor (Kranner y col., 2010; Li y Pritchard, 2009). Sin embargo, el resultado más novedoso y destacable es que la mortalidad de estas plántulas se redujo considerablemente en las semillas conservadas en frío. La patogenicidad es un factor determinante en el reclutamiento de plántulas (Kranner y col., ), siendo una de las mayores causas de mortalidad de éstas durante su primer mes de vida en los bosques tropicales (Comita y col., 2014).
En estos bosques, el ataque de los patógenos suele tener efectos más agudos sobre aquellas plántulas que crecen en sombra (Augspurger y Kelly, 1984), como es el caso de C. platanifolia. La conservación en frío, por tanto, no sólo mantendría el vigor de las semillas, sino que también reduciría la actividad de los patógenos, limitando su impacto negativo sobre la fisiología de las semillas y el establecimiento de las plántulas. Es común en los bosques tropicales que la mortalidad de las plántulas debido a la acción patogénica también esté ligado a su densidad (Alvarez-Loayza y Terborgh, 2011; Augspurger y Kelly, 1984). Nuestra metodología muestra que pueden obtenerse bajas tasas de mortalidad aun realizando la siembra a altas densidades (∼ 83 % de supervivencia en semillas conservadas en frío, habiendo 2 cm de distancia entre plántulas).
Conclusiones
Consideramos que la conservación en frío pudo disminuir la mortalidad tanto limitando el debilitamiento causado por el envejecimiento como ralentizando el crecimiento de los patógenos, conllevando a una menor probabilidad de infección letal. La fácilmente replicable y económica metodología de vivero utilizada en este estudio puede ser empleada para la producción de plántulas en los planes de reforestación de C. platanifolia. De esta forma, el vivero puede ser ocupado en diferentes tiempos de sembrado, planificando la activación y siembra de la semilla mantenida en frío según conveniencia, optimizando así la eficiencia en la producción de plántulas.
Esta metodología podría emplearse también en otras especies de Malváceas-Bombacáceas, pues casi la mitad de las especies de esta familia están catalogadas como al menos vulnerables (IUCN, 2019). La metodología podría igualmente testarse en otras especies importantes y amenazadas del ChocóDarién con semillas de características similares, entre las que se encuentran Quercus humboldtii Bonpl. (Fagaceae), Prioria copaifera Griseb. (Fabaceae), Caryocar amygdaliferum Mutis (Caryocaraceae) y Anacardium excelsum (Bertero Balb. ex Kunth) Skeels (Anacardiaceae).

