











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
I. INTRODUCTION
According to six major international temperature datasets consolidated by the World Meteorological Organization (1), the last eight years have been the warmest on record globally. In 2022, the global average temperature exceeded pre-industrial levels (1850-1900) by approximately 1.15°C (ranging from 1.02°C to 1.27°C). These data are a cause for growing concern, especially considering the commitment established in the Paris Agreement to limit global warming to significantly below 2°C compared to pre-industrial levels and to strive for a 1.5°C limit by the year 2100. Unfortunately, 2022 marked the eighth consecutive year in which global annual temperatures have exceeded pre-industrial levels by at least 1°C, representing a significant challenge in achieving these crucial goals.
Forests play a crucial role in reducing greenhouse gas emissions (GHG) and mitigating climate change, as established by the United Nations Framework Convention on Climate Change (UNFCCC) (2). Globally, forests store approximately 861 gigatons (Gt) of carbon, with 44 % in the soil, 42 % in live biomass, 8 % in dead wood, and 5 % in litter (equivalent to nearly a century of annual fossil fuel emissions). According to the World Bank (3), the carbon sink benefits of forests are diminished by deforestation and degradation, with 80 % of global deforestation being driven by agriculture. Between 2001 and 2021, emissions from deforestation and other forest disturbances were 8.8 gigatons Gt CO2e per year, while forest absorptions were -16.6 Gt CO2e per year, resulting in an average net sink of -7.7 Gt CO2e per year (4). Forest biomass provides estimates of carbon stocks in forest vegetation, as approximately 50 % of it is composed of carbon. Over the last 22 years, forest absorptions in all climatic domains (tropical, subtropical, temperate, boreal) have exceeded emissions from forest disturbances (5).
In 2010, Ecuador had approximately 19.1 million hectares of natural forest, representing around 75 % of its total land area. However, by 2021, the country experienced a loss of 29.7 thousand hectares of natural forest, equivalent to an estimated release of 21.5 million tons of CO2 into the atmosphere (6). Four provinces (Sucumbíos, Orellana, Esmeraldas and Manabí) collectively accounted for 55 % of all tree cover loss during this period (7). Taking Manabí as an example, it had 1.16 thousand hectares of natural forest at the beginning of the analyzed period, representing approximately 60 % of its territory. Unfortunately, by 2021, this province lost 473 hectares of natural forest, which is equivalent to the release of approximately 262 thousand tons of CO2 (8). Additionally, during the same period, Portoviejo also experienced significant tree cover loss. In 2010, the canton had 50.4 thousand hectares of forest, covering approximately 53 % of its land area. However, by 2022, the loss amounted to 9.46 hectares, with an estimated release of 5.01 kilotons of CO2 into the atmosphere (9). Currently, Ecuador hosts approximately 12.6 million hectares of native forest, representing around 50.73 % of its national territory. These data underscore the urgent need to take concrete measures to preserve and protect natural resources, as well as to comprehensively and effectively address the challenges associated with climate change in the country.
In the specific case of Ecuador, it was estimated that in the year 2000, the density of aboveground woody biomass was 253 tons per hectare, with a total aboveground biomass (AGB) of 4.54 gigatons. Subsequently, between 2001 and 2022, the country’s forests showed annual emissions of 27.6 megatons of CO2e, while carbon removals reached -63.1 megatons of CO2e per year. This represents a net carbon sink of -35.5 megatons of CO2e per year (6). These data are fundamental to understanding the role of forests as regulators of the carbon balance and their contribution to mitigating climate change (10).
Forest biomass is essential for estimating carbon stocks in forest vegetation and assessing changes in forest structure. The total biomass is derived from the difference between production through photosynthesis and consumption through respiration, making it a valuable measure to understand the state of forests (11). However, it is important to highlight that forest biomass depends on its composition and structure. Therefore, preserving the integrity of forests and protecting them from deforestation and degradation is crucial to ensure their function as carbon sinks and make a significant contribution to the fight against climate change.
The main objective of this study was to determine the composition and structure of the vegetation, examining species interactions, and estimating carbon stored in its AGB using an allometric equation proposed for mixed dry forests. The resulting analysis intends to discuss the importance of the composition and structure of the ecosystem in terms of the carbon it can store. Subsequently, the results could serve as an important input for decision making for the conservation of tree species in the study area.
II. METHODOLOGY
The study area is located in the forestal citadel at the southeastern of Portoviejo canton, in the province of Manabí, situated at coordinates -1.0187576 and -80.455844 (Figure 1). Field research was conducted during the period between July and August 2022. For data collection, 10 sampling plots measuring 10 x 20 m were established, resulting in a total area of 2 000 m2. In each plot, height and diameter at breast height (DBH) measurements were recorded for all trees with a DBH equal to or greater than 5 cm.
The taxonomic identification of species was carried out in two phases. Initially, valuable assistance was provided by local experts from the Jardín Botánico of the Universidad Técnica de Manabí (UTM), who contributed with their knowledge and experience to identifying the species present in the study area. Additionally, international databases such as WFO (The World Flora Online http://www.worldfloraonline.org/), International Plant Name Index (IPNI), Nomenclator from the Missouri Botanical Garden, and VAST (VAScular Trópicos / https://www.tropicos.org/) were used as complementary sources. These platforms provided precise and updated taxonomic information, ensuring rigor and accuracy in the classification of the studied species.
A. Composition and structure
A structural study was conducted based on taxonomic data encompassing four fundamental parameters to understand the ecology of the vegetation community:
Abundance: was analyzed the abundance of individuals per species in the sample. Were considered two types of abundance: absolute abundance (Aba), which represents the total number of individuals per species, and relative abundance (Abr), which shows the proportion of individuals of a species relative to the total number of individuals found in the study area.
Frequency: was determined the number of plots in which each species appears, evaluating both absolute frequency (Fra) and relative frequency (Frr). Relative frequency was calculated considering the number of plots in which a species appears in relation to the total number of inventoried plots.
Dominance: this parameter is related to the degree of coverage of species and was analyzed for both absolute dominance (Doa), which measures the basal area occupied by each species, and relative dominance (Dor), which shows the proportion of the absolute dominance of a species in relation to the absolute dominance of all species present.
Importance Value Index (IVI): proposed by Curtis & McIntosh (12), this index was calculated for each species by summing the relative abundance, relative frequency, and relative dominance. The IVI allows for comparing the ecological importance of each species within the forest, offering a comprehensive view of the composition, structure, site quality, and dynamics of the study area.
The exhaustive analysis of these parameters provided a complete understanding of the vegetation community in the studied forest. The evaluation of the IVI facilitated the understanding of the interaction between different species and their role in the ecological balance of the studied area.
Subsequently, using the absolute abundances of different species in each plot as input, a cluster analysis based on the Jaccard index was used to group the plots according to the similarity of their communities. From the obtained groups, the IndVal% was calculated along with the corresponding p-values for each species in each group of plots. IndVal% was used to measure the association of each species with a specific group, while p-values assessed the statistical significance of these associations.
Additionally, a diameter analysis was performed using 10 diameter classes with an interval of 18 cm to determine the structure of the forest under study.
B. Carbon stored in AGB
The allometric equation proposed by (13) for mixed dry forests was applied to estimate the AGB of trees based on dendrometric variables. The variables used to feed the model were represented by total height (ht) in meters, diameter at breast height (DBH) in centimeters, and wood density (ρ) in grams per cubic centimeter. To obtain wood density was used information from the “Global wood density database” (14). In cases where the data was not available was used the density of the genus, family (15), or an average of the individuals present in the plot.
Once the estimated AGB in kilograms of dry matter per tree was calculated, the total biomass per hectare (Mg ha-1) was determined. To achieve this, the biomass value obtained per plot was multiplied by a conversion factor that depended on the size of the plot. For the conversion of AGB to carbon were followed the guidelines established by the Intergovernmental Panel on Climate Change (IPCC) (16). It was assumed that the carbon content in the AGB of each living tree is equivalent to 50 % of the AGB (17). Thus, the amount of carbon in the AGB was obtained in megagrams per hectare (Mg C ha-1).
Once the data was collected, the Kruskal-Wallis test was used for the statistical analysis of the carbon values found per plot. This non-parametric test was chosen for its suitability to compare the means of three or more independent groups without assuming a normal distribution in the data. The Kruskal-Wallis test evaluated the null hypothesis that there are no significant differences in the amount of carbon stored among the plots of the dry forest. A typical significance level of 0.05 was established for the test.
III. RESULTS
A. Composition and structure
A total of 148 individuals belonging to 21 species within 19 genera and 12 families were identified (Table I). The Fabaceae family was the most diverse, with 44 individuals, followed by Capparaceae with 32 individuals, and Malvaceae with 25 individuals. Regarding abundance, it was determined that C. Flexuosa exhibited the highest absolute abundance (28 individuals) and relative abundance (19.18 %). This indicates that this species is dominant in terms of the number of individuals present in the study area. The species E. Ruizii, C. Flexuosa and A. Macracantha showed high relative frequency, suggesting that they are widely distributed in various plots within the dry forest area. On the other hand, C. Trischistandra displayed the highest absolute dominance (4.96 m² ha-1) and relative dominance (66.38 %), indicating that this species occupies a significant space within the forest and may have an important influence on the structure of the vegetation community. Furthermore, C. Trischistandra also stood out with the highest IVI value (77.39), suggesting that it plays a crucial role in the ecological balance of the dry forest.
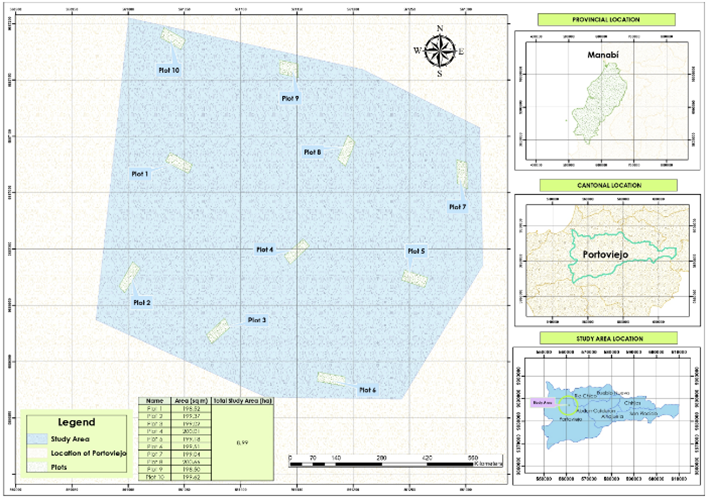
Fig. 2 Clustering of plots according to Jaccard index based on species similarity. Where: G1 = Group 1, G2 = Group 2, G3 = Group 3, G4 = Group 4.
Additionally, the speciesE. RuiziiandC. Flexuosaalso exhibited considerable IVI values (36.87 and 33.54, respectively).
The clusters obtained through the cluster analysis and Jaccard index (Figure 2) result from the classification of the 10 plots based on the similarity of their species communities.
Group G1 consisted of:
A. Macracantha
C. Inermis
C. Vitifolium
E. Ruizii
G. Spinosa
M. Acantholoba
M. Glabra
Group G2 included:
A. Macracantha
A. Pubescens
B. Aculeata
C. Flexuosa
C. Lutea
C. Scabrida
C. Scabridum
E. Ruizii
P. Dulce
Z. Thyrsiflora
Group G3 comprised:
A. Macracantha
A. Pubescens
B. Aculeata
B. Graveolens
B. Sprucei
C. Coriaria
C. Flexuosa
C. Ruiziana
C. Vitifolium
C. Trischistandra
E. Ruizii
P. Dulce
V. Parviflora
Z. Thyrsiflora
Lastly, Group G4 consisted of:
A. Macracantha
B. Sprucei
C. Flexuosa
C. Inermis
M. Acantholoba
Z. Thyrsiflora
Table II Indicator Value Index (IndVal%) along with corresponding p-values for each species in each group of plots
Certain species showed significantly high IndVal% values in specific groups of plots, suggesting a strong association with those specific communities (Table II). In this regard, it was observed that althoughA. Macracanthadoes not exhibit very high values, it is associated with the other species present in all groups, with a predominant tendency towards Group 4.C. Inermis, G. Spinosa, andM. Glabradisplayed IndVal% values of 100 % in Group 1, demonstrating that they can be considered indicators of those particular communities.
The speciesC. TrischistandraandB. Spruceishowed interesting patterns regarding their association with different groups of plots in the dry forest. On one hand, the IndVal% for C.Trischistandrain Group 3 was 100 %, indicating that it is highly represented in this group and shows a strong association with the other species present in it (p < 0.05). On the other hand,B. Spruceiexhibited a similar behavior in terms of IndVal% (66.67 %) and p-value (0.037). In both cases, considering the p-value, it can be determined that these associations are not random and are statistically significant. This suggests that both species may have specific characteristics that make them prefer the environmental conditions and coexisting species in each of the mentioned groups.
The diametric analysis resulted in a reverse J-shaped distribution (Figure 3)(18). This distribution refers to the configuration of the diameter structure of a forest, where there is a predominance of young and small-diameter individuals, followed by a marked decrease in density in intermediate diameters, and finally, an increase in the number of individuals as the diameter increases. This curve represents a typical pattern of natural regeneration and development of the forest community(19).
The origin of this distribution can be influenced by various factors, such as the history of natural or anthropogenic disturbances(20), the interaction between different strata of the forest and the regeneration capacity of the present species(21). In any case, this type of distribution can indicate efficient biological functionality, strong recruitment capacity, a healthy state and a stable population(22).
In any case, the young individuals, which make up the lower diameter classes, are often the product of recent regeneration or succession events(23), which may be associated with selective logging or biotic factors. These conditions create open spaces where certain tree species establish and compete for resources, leading to a high density of small-sized young individuals(24).
As these trees mature and survive competition, they grow and become the individuals that form the wider part of the reverse J-shaped curve, i.e., the trees with intermediate diameters. At this stage, the process of self-thinning and interspecific competition becomes more noticeable, resulting in a thinning of tree density(25). Some weaker trees may die or be outcompeted by more vigorous individuals adapted to the environment, contributing to the reduction of density in this zone of the curve.
Finally, in the right side of the reverse J-shaped curve, we find the largest and oldest trees. These individuals have overcome the earlier stages of development, withstood disturbances, and successfully competed for limited resources. Their presence indicates greater stability and longevity within the forest ecosystem(26).
Understanding the reverse J-shaped diametric distribution is essential for making informed decisions in sustainable forest management(27). By analyzing this curve, forest managers can assess the regeneration status, evaluate the structure and health of the forest, and plan management strategies that promote proper regeneration, optimal growth of desirable species, and conservation of natural resources. Moreover, this knowledge enables addressing challenges such as the conservation of threatened species, mitigation of the effects of climate change and promotion of forest resilience to future disturbances(28).
B. Carbon stored in AGB
The Kruskal-Wallis test did not yield sufficient evidence to claim the existence of statistically significant differences in the amount of carbon stored in the different plots of the dry forest. However, it was possible to observe that certain species store more carbon than others, primarily because they are more abundant in the plots, have larger diameters, heights or wood density (Figure 4). In this regard, it was determined that the study area can store 70.47 Mg C ha-1 in AGB. These values are somewhat higher than those found by the Ministry of the Environment (29) and (30) possibly due to the composition and structure of the forest.
V. CONCLUSION
The results of the structural analysis in this study provide a detailed and revealing insight into the composition and structure of this ecosystem. Species with higher abundance, frequency, and dominance - as well as those with significant IndVal% values- are of particular relevance for understanding the dynamics and ecology of the dry forest. These findings help identify which species play key roles in the community and which may serve as indicators of specific conditions.
It is essential to note that the analysis of species clusters should consider not only the structural parameters of the forest but also relevant environmental factors. These factors can influence the distribution and coexistence of species, and their consideration is vital to fully comprehend the observed clustering patterns. The obtained results may vary based on spatial scale and study area, necessitating detailed analyses in different sites to gain a more comprehensive understanding of the variability in the plant community of the dry forest.
In this regard, the results of this study may have significant implications for estimating carbon stored in live AGB in the dry forest. Species with high IndVal% values and abundance could be significant contributors to the total AGB of the ecosystem. Therefore, the conservation and effective management of these species could have a positive impact on carbon capture and storage, which is relevant in the context of climate change and efforts to mitigate the effects of global warming.

