











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Forma sugerida de citar:
Cedeño-Zambrano, J., García-Párraga, J., Solórzano-Cobeña, C., Jiménez-Flores, L., Ulloa-Cortazar, S., López-Mejía, F., Avellán-Vásquez, L., Bracho-Bravo, B., y Sánchez Urdaneta, A. (2022). Fertilización con magnesio en plátano ’Barraganete’ (Musa AAB) Ecuador. La Granja: Revista de Ciencias de la Vida. Vol. 35(1):8-19. http://doi.org/10. 17163/lgr.n35.2022.01.
1. Introducción
La producción mundial de plátano (Musa sp.) durante los años 2012-2017 se distribuyó en África (60,60%), América (26,50%), Asia (12,80%) y Oceanía (0,10%), lo cual es un indicativo de la importancia económica y alimenticia de este cultivo para la población (FAOSTAT, 2018). En América, Asia y África predominan temperaturas y humedad relativa altas en las zonas subtropicales y tropicales de estos continentes, los plátanos constituyen un cultivo de relevancia importancia por el área sembrada que representan (Ramos y col., 2016).
De acuerdo con el Centro de Estudios para el Desarrollo Rural Sostenible y la Soberanía Alimentaria (CEDRSSA, 2019), la India es el principal país productor de plátano con 30 477 000 t, 2,67 veces mayor a China (11 422 956 t), su más cercano competidor; señalando además que en Latinoamérica hay seis países que destacan en producción de plátano, tales como Brasil (6 675 100 t), Ecuador (6 282 105 t), Guatemala (3 887 439 t), Colombia (3 786 672 t), Costa Rica (2 552 822 t) y México (2 229 519 t).
En Ecuador, el plátano es un rubro de exportación y una fuente de empleo en diversas zonas del país. Debido a la importancia de este cultivo, se hace necesario generar herramientas confiables para que el agricultor maneje el cultivo de una manera adecuada y rentable (Tumbaco y col., 2015). Además de su importancia por el aporte que este genera para la socioeconomía y seguridad agroalimentaria, fortaleciendo el empleo directo (mano de obra fija) e indirecto (mano de obra ocasional, valor agregado de productos) y suministrando carbohidratos a la mayor parte de la población. De acuerdo con el Instituto Nacional de Estadística y Censo (INEC, 2020) en Ecuador en el 2019 hubo 115 069 ha sembradas de plátano como monocultivo y 45 194 ha asociadas con otros cultivos, la producción correspondió a 582 706 y 166 745 Tm, equivalente a 5 064 y 3 690 kg·ha−1, respectivamente.
Las exportaciones de banano y plátano en Ecuador correspondieron a $3,27 miles de millones, representando 17,5% del total de las exportaciones no petroleras (OEC, 2017; Álvarez y col., 2020); no obstante, el Banco Central de Ecuador (BCE, 2020) señaló que la exportación de plátanos en Ecuador para 2019 correspondió a 211 732,6 Tm. En un estudio realizado por Molero y col. (2008), la extracción de nutrientes por el cultivo de plátano cv. Hartón al momento de la cosecha fueron aproximadamente de N: 150; P: 60; Ca: 215; Mg: 140; Mn: 12; Fe: 5; Zn: 1,5; B: 1,25 y Cu: 0,5 kg·ha−1·año−1; por lo que resultó importante utilizar dosis de restitución para mantener la fertilidad del suelo y garantizar una alta producción. Estos mismos autores han señalado que el nitrógeno, el potasio y en menor cantidad el magnesio en el cultivo de plátano son los elementos más importantes para su crecimiento y producción.
En contraposición a esto, Cobeña y col. (2020) encontraron que en plátano ‘Barraganete’ los elementos nutritivos que fueron extraídos en mayor cantidad fueron K, Ca, N, P y Mg, cuando se realizaron fertilizaciones con N y K2O; con respecto al contenido de P y Mg, y no hubo un comportamiento mantenido de extracción de estos dos elementos. Avellán y col. (2020) evaluaron la exportación de P en plátano ‘Barraganete’, encontrando que este presentó movilidad baja en el suelo, y absorbiendo el que se encontró alrededor de las raíces; lo que indica que al realizar la fertilización, el P estuvo disponible y en forma soluble, pero fue fijado casi de manera inmediata, dejando de estar aprovechable para la planta; por lo que el fraccionamiento del P fue sugerido como una alternativa para el incremento de su eficiencia.
Por otro lado, el Mg tiene gran importancia en las plantas. El 75% de la cantidad de Mg contenido en las hojas participó en la síntesis de proteínas y entre el 15 y 20% del Mg total estuvo asociado con los pigmentos clorofílicos (White y Broadley, 2009), interviniendo como cofactor de una serie de enzimas involucradas en el metabolismo y fijación del carbono fotosintético (Hermans y col., 2013).
Entre el 90 y 98% del Mg presente en el suelo no se encuentra disponible para ser absorbido por las plantas, sino que está incorporado en la estructura cristalina de los minerales (Senbayram y col., 2015). La forma aprovechable del Mg para ser absorbido por las plantas es como ión Mg2+, el cual posee el radio iónico más pequeño, pero el radio hidratado más grande entre los cationes (Maguire y Cowan, 2002). Esto hace que se una débilmente a los coloides del suelo cargados negativamente y a las paredes de las células de las raíces, lo que genera que el Mg se pierda fácilmente (Grzebisz, 2011). Al mismo tiempo, la fertilización excesiva con K+ y NH+ 4 es antagónica con la absorción de Mg, incrementando su deficiencia. La deficiencia de Mg aumenta en los suelos ácidos saturados con cationes de H+, Al3+ y Mn2+, donde ocurre lixiviación intensiva, especialmente en áreas con altas precipitaciones (Gransee y Führs, 2013).
Aun ante la importancia del Mg2+ en el crecimiento y desarrollo de las plantas, el mecanismo molecular de las células vegetales que regulan la homeostasis del Mg2+, así como los mecanismos moleculares relacionados con el transporte de magnesio, son poco conocidos. Se requiere de Mg2+ para que se presente un adecuado funcionamiento de las numerosas actividades celulares relacionadas con la síntesis de clorofila, por ser el componente principal de la clorofila, la estabilidad de la membrana y la activación enzimática (Guo y col., 2016).
Por otro lado, la nutrición con Mg en las plantas debe considerarse como un importante problema mundial, no solo en lo que se refiere a la seguridad alimentaria, sino también para la salud humana, ya que el Mg es un nutriente esencial en la alimentación humana y animal, y las plantas constituyen la principal fuente de ingesta diaria de este elemento. En este sentido, se han realizado experimentos donde se ha evaluado el efecto del magnesio en plantas; no obstante, esta información en plátano es escasa, particularmente en Ecuador. Por lo tanto, el objetivo es evaluar la respuesta a la fertilización con magnesio en el cultivo de plátano ‘Barraganete’ (Musa AAB), en el cantón El Carmen, Manabí, Ecuador.
2. Materiales y Métodos
La investigación se realizó entre 2018-2019, en la Granja Experimental Río Suma, Universidad Laica “Eloy Alfaro” de Manabí, extensión El Carmen, geográficamente ubicada en la Provincia de Manabí, Cantón El Carmen, en coordenadas UTM (−0,259503 S; −79,427558 0), en clima trópico húmedo. Las características agroecológicas correspondieron a altitud 260 msnm, temperatura de 24,15 °C, precipitación anual de 2 684 mm, humedad relativa de 85,6%, heliofanía de 553 horas·luz−1·año−1 y evaporación de 1 064,30 mm·año−1 (Climatedata.org, 2019).
El análisis de suelo del área de estudio indicó niveles medios de MO (4,48%), C.E. no salino (0,08 ds·m−1), pH de 5,7 (medianamente ácido), contenidos bajos de NH4 (11,61 ppm), P (4,56 ppm), S (2,14 ppm), Mg (0,90 meq·100 g−1), sumatoria de bases 8,40 meq·100 g−1, Mn (9,70 ppm), y relación Mg/K (1,80), Ca+ Mg/K (15,80). Niveles altos de K (0,50 meq·100 g−1), Ca (7,00 meq·100 g−1), Cu (5,80 ppm), Fe (123,10 ppm), Zn (23,20 ppm) y relación Ca/Mg (7,78). La textura es franco-arenosa (62% de arena, 28% de limo y 10% de arcilla) (Avellán y col., 2020). Después que el cultivo alcanzó la edad media se tomaron muestras de suelo y del área foliar al final del ciclo del cultivo. Las muestras de suelo fueron analizadas en el Laboratorio Agrolab, adscrito a la Red de Laboratorios de Agua y Suelos de Ecuador (Relase).
La investigación se evaluó en una plantación de plátano ‘Barraganete’ (Musa sp. AAB) de primer ciclo, con una distancia de siembra de 2,5 m entre hileras × 1,8 m entre plantas (4,5 m2), para un total de 2 222 plantas·ha−1, correspondiendo a un manejo de alta densidad de plantas. Se incluyeron 288 plantas en la investigación, y se distribuyeron en tres bloques. Se realizaron aplicaciones de fertilizante fraccionados en tres dosis (N-P-K y MgO). Los tratamientos siguieron un diseño experimental en bloques totalmente al azar, con tres repeticiones.
Los tratamientos fueron fertilizados con MgO a seis niveles (0, 25, 50, 75, 100 y 125 kg·ha−1) y una dosis estándar de 80 kg·ha−1 de N, 40 kg·ha−1 de P2O5 y 150 kg·ha−1 de K2O, fraccionados en tres partes iguales y aplicado al suelo cuando la planta emitió las hojas 12, 16 y 20. Los fertilizantes comerciales que se utilizaron fueron urea con 46% de N, fosfato diamónico (DAP) con 18% de N y 46% de P2O5, muriato de potasio con 60% de K2O y óxido de magnesio con 30% de MgO.
La unidad experimental estuvo conformada por 16 plantas, de allí se seleccionaron cuatro plantas de la parte central para evitar el efecto de bordura y realizar sobre ellas las evaluaciones. Las plantas fueron trasplantadas al emitir la hoja cinco. Los promedios de los tratamientos se compararon mediante la prueba de Tukey (P<0,05). Para el procesamiento de los datos se utilizó el procedimiento GLM del programa SAS®, versión 15.1 (Statistical Analysis System, 2020).
Las variables morfo-fisiológicas (altura de la planta, perímetro del pseudotallo, número de hojas, largo y ancho de la hoja 3 y área foliar), fueron evaluadas a través del tiempo con mediciones cada 8 días, por lo que se analizó utilizando la metodología de medidas repetidas en el tiempo a través del procedimiento MIXED (Statistical Analysis System, 2020); de igual forma, se seleccionaron modelos polinómicos de segundo grado que explicaron mejor el comportamiento a través del tiempo de las variables de crecimiento. La representación gráfica de las variables se realizó con el software SigmaPlot. Además, se tomaron variables relacionadas con el rendimiento (número de manos, número de dedos, longitud de dedos, diámetro de los dedos exportables, biomasa del fruto y del racimo, rendimiento por hectárea, concentración edáfica y foliar de Mg, concentración de Mg en el fruto, exportación de Mg en el fruto, eficiencia agronómica y factor parcial de productividad). El área foliar se determinó a través de las medidas obtenidas de la longitud y ancho de la tercera hoja y se calculó con la siguiente fórmula:
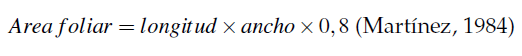
Para las variables relacionadas con la eficiencia de la fertilización se determinaron la exportación de Mg, para lo cual se consideró el porcentaje de biomasa seca del fruto multiplicado por el rendimiento del cultivo por hectárea y multiplicado por el porcentaje de concentración de magnesio; de esta manera se calculó la cantidad de magnesio que salió del cultivo mediante el fruto y la respuesta se obtuvo en kg·ha−1.
La eficiencia agronómica (EA) se definió como el aumento del rendimiento a partir de la fertilización por unidad de nutriente aplicado (Dobermann, 2007) y se calculó considerando el rendimiento de las parcelas fertilizadas (RPF) menos el rendimiento de la parcela testigo (RPT), dividido entre la dosis aplicada (DA), se expresó en kg de fruto·kg−1 de nutriente aplicado.
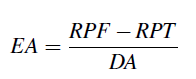
El factor parcial de productividad permite evaluar la eficiencia de un nutriente para la producción (Bruulsema y col., 2011). Se calculó con la fórmula FPP =R /D en el cual R = rendimiento de la parcela cosechada (kg·ha−1) y D = dosis de nutriente aplicado (kg·ha−1) (Dobermann, 2007).
3. Resultados y Discusión
3.1 Altura de la planta y perímetro del pseudotallo
Se encontraron diferencias significativas (P<0,01) para esta variable de manera general por efecto de los tratamientos para los muestreos realizados en la semana 10 (T25-T50; T75-T125 y T100-T125), y a partir de la semana 11 a la 53 las diferencias se presentaron entre T0-T75, T0-T100, T0-T125, T50-T75 y T50-T100. Para las semanas indicadas, T25 fue el que presentó la mayor altura de la planta, alcanzando 3,47 m a las 53 semanas después de la siembra; el T0 fue superior a T75 y T100.
La altura de las plantas tuvo la misma tendencia para todos los tratamientos a través del tiempo, ajustándose a un polinomio de segundo grado (Y= a + bx + cx2), e indicando que la fertilización con magnesio incrementó la longitud de las plantas de plátano ‘Barraganete’ con T50, T125 y T0 comparado con T75 y T100; estos dos últimos presentaron valores similares a T25 hasta la semana 46, a partir de allí se incrementó hasta valores similares a T0, T50 y T125 (Figura 1). Este comportamiento polinómico de segundo orden sugiere que al acercarse la emergencia de la inflorescencia se desacelera el crecimiento de la planta hasta que alcanza su altura final.
La altura promedio obtenida en esta investigación a las 53 semanas fue de 3,47 m, similar a la reportada por Cayón y col. (2004) que estuvo entre 3,2−3,4 m al evaluar densidades y arreglos de siembra en un rango de 1 500 a 3 000 plantas·ha−1, sin diferencias estadísticas entre ellas.
Se encontraron diferencias significativas (P<0,01) para el perímetro del pseudotallo por efecto de los tratamientos para todas las semanas donde se realizaron las evaluaciones. Hubo diferencias en la semana 10 entre T0-T25; T0-T50 y T100-T125 y entre las semanas 11 a la 53 en T0-T75, T0-T100, T0-T125, T25-T100, T25-T125, T50-T100 y T50-T125. Esta variable a través del tiempo se ajustó a un polinomio de segundo orden (Y= a + bx + cx2). Hubo una tendencia similar entre los tratamientos a través del tiempo, indicando que la fertilización con MgO afectó el diámetro del pseudotallo del plátano ‘Barraganete’ (Figura 2).
En este sentido, se reconoce la importancia del Mg en diversos procesos metabólicos y reacciones en las plantas, particularmente en la fotosíntesis y en el contenido de clorofila; por ello, Cakmak y Yazici (2010) señalaron que las plantas que presentan deficiencias de Mg acumularon hasta cuatro veces más sacarosa al compararlas con las que presentaron un adecuado contenido de Mg, generando una severa inhibición del transporte de sacarosa en las hojas hacia otros órganos receptores de la planta (raíces, tallos, entre otros). Una adecuada nutrición con Mg durante los periodos de transporte de carbohidratos de las hojas a las células en otros sitios de la planta garantiza maximizar el transporte de carbohidratos hacia los órganos receptores, con lo cual se promovió el crecimiento y altos rendimientos.
El promedio general del perímetro del pseudotallo fue de 52,23 cm, similar al indicado por Barrera, Combatt y Ramírez (2011) de 52,23 cm en promedio para el primer ciclo de producción y de 48,15 cm para el segundo ciclo en plátano Hartón con una densidad de 1 111 plantas·ha−1; no obstante, con la aplicación de micorrizas + lombriabono fue de 60,08 y 41,5 cm, para cada ciclo de producción, respectivamente, siendo menor que el obtenido por Pinchao (2018) al evaluar en plátano ‘Barraganete’ el efecto de diferentes niveles de K2O y MgO, obteniendo en promedio 70 cm en esta misma variable con una densidad de 2 222 plantas·ha−1.

Figura 1. Altura de las plantas de plátano ‘Barraganete’ (Musa ABB) bajo diferentes dosis de fertilización con MgO, en El Carmen, Ecuador. Datos recolectados cada 8 días. Se presentan los valores estimados. T0= testigo (0 kg·ha−1 de MgO, negro), T25= 25 kg·ha−1 de MgO (rojo), T50= 50 kg·ha−1 de MgO (verde), T75= 75 kg·ha−1 de MgO (amarillo), T100= 100 kg·ha−1 de MgO (azul) y T125= 125 kg·ha−1 de MgO (magenta).
3.2 Ancho, largo y área foliar
Se encontraron diferencias significativas (P<0,01) para el ancho de la hoja en la semana 10 (T0-T25, T0T75, T0-T100, T0-T125, T25-T50, T25-T125, T50-T75, T50-T100, T50-T125 y T75-T100; para la semana 11 (T0-T25, T0-T100, T25-T50, T25-T75, T25-T125, T50- T100, T75-T100 y T100-125; y a partir de la semana 12 hasta la 53 entre T0-T25, T0-T100, T25-T50, T25T75, T25-T150, T50-T100, T75-T100 y T100-T125 por efecto de los tratamientos.
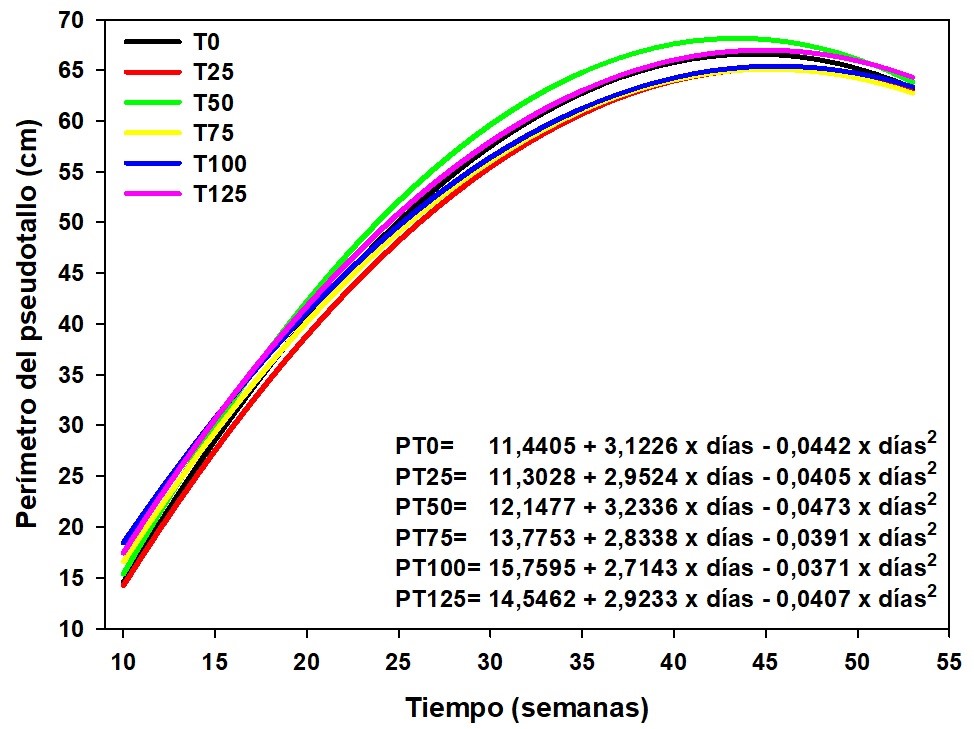
Figura 2. Perímetro de pseudotallo de plátano ‘Barraganete’ (Musa ABB) bajo diferentes dosis de fertilización con MgO, en El Carmen, Ecuador. Datos tomados cada 8 días. Se presentan los valores estimados. T0= testigo (0 kg.ha-1 de MgO, negro), T25=25 kg.ha-1 de MgO (rojo), T50= 50 kg.ha-1 de MgO (verde), T75= 75 kg.ha-1 de MgO (amarillo), T100= 100 kg.ha-1 de MgO (azul) y T125= 125 kg.ha-1 de MgO (magenta).
Igualmente hubo diferencias estadísticas (P<0,01) para el largo de la hoja durante todo el periodo evaluado entre T0-T25, T25-T50, T25-T75, T25-T125, T50-T100, T75-T100 y T75-T125. El área foliar presentó diferencias estadísticas en la semana 10 entre T0-T25, T0-T50, T0-T100, T50-T100, T75T100 y T75-T125; y desde la semana 11 a la 53 entre T0-T25 y entre T25 con T50, T75, T100 y T125.
Los resultados indicaron que la fertilización con MgO influyó en el desarrollo de las hojas. Tanto el ancho de las hojas como el área foliar a través del tiempo presentaron una tendencia similar, donde T0 fue el que tuvo menor desarrollo de las hojas (Figura 3). Los tratamientos tuvieron un comportamiento polinómico de segundo grado (Y= a + bx + cx2) para estas variables, donde a partir aproximadamente de la semana 30 del desarrollo del cultivo hubo una tendencia a incrementarse el ancho de las hojas entre los diferentes tratamientos, a excepción de T0 que se mantuvo a través del tiempo.
Con la dosis de 125 kg·ha−1 hubo un incremento en el ancho de la hoja de 1,12 veces con respecto a T0, para la semana 53. En promedio T0 presentó un ancho de 0,68 m, mientras que T50 y T125 tuvieron en promedio 0,72 m. El desarrollo de las hojas tuvo similar comportamiento a lo señalado por Aristizábal (2008), quien indicó que las hojas terminales disminuyeron su tamaño en las últimas semanas antes de la emisión de la inflorescencia (bellota), y en esta investigación se realizó sobre la hoja 3 de la planta.
Con respecto al largo de la lámina foliar, en la Figura 3 se evidenció que a partir de la semana 15 y hasta la semana 30 se observó un crecimiento más acelerado de las hojas, y en promedio fue de 1,88 veces mayor en el rango indicado. A partir de la semana 30 y hasta la semana 53 el crecimiento de las hojas fue más lento (incremento de 1,04 veces), lo cual evidenció que en las últimas semanas la longitud de las hojas tuvo una pendiente menor, siendo casi horizontal el incremento del crecimiento. Tanto la anchura como la longitud de las hojas fue evaluada en la hoja 3, la cual ha venido siendo utilizada como hoja de referencia en el cultivo.
De acuerdo con Martínez y Cayón (2011) la fase logarítmica (fase vegetativa) fue de lento crecimiento, seguida de un cambio progresivo en su tasa de velocidad, evidenciado por la pendiente de la curva, y con incremento exponencial de la anchura y longitud de las hojas; la diferenciación marcó la terminación de esta fase. Continuó la fase lineal (fase vegetativa-reproductiva), presentándose en la planta crecimiento acelerado y constante, y por ende con estabilidad de la pendiente de la curva; en esta fase se destacó el desarrollo y elongación del tallo floral, y culminó con la emergencia de la inflorescencia (floración). El ciclo del cultivo se completó al iniciar la fase de senescencia (fase reproductiva-productiva), con la floración y posterior desarrollo del racimo; el ritmo de crecimiento de la planta disminuyó ya que se movilizaron los asimilados de hojas y pseudotallo (fuentes) al fruto (sumidero); coincidiendo este comportamiento con el obtenido en esta investigación.
La dinámica del área foliar a través del tiempo sugirió que el área foliar tendió a disminuir conforme la planta se aproximaba a la emisión de la inflorescencia, lo cual podría deberse a una menor tasa de emisión de hojas; disminución del tamaño de la lámina foliar, menor longevidad de las últimas hojas producidas, o al efecto combinado de estos hechos.
En la Figura 3 se observa que T0 presentó un comportamiento más uniforme que las plantas fertilizadas, con la menor área foliar al final del periodo de investigación, el cual fue 1,19 veces menor que T125. Esto debido a que la carencia de Mg inhibe el crecimiento de las plantas, acelera el proceso de envejecimiento y genera pérdidas tanto en la producción como en la calidad de los cultivos (Verbruggen y Hermans, 2013), ya que está implicado en el proceso de fotosíntesis, por lo que su deficiencia disminuyó la tasa fotosintética y bloqueó la retribución de carbohidratos a los órganos demandantes, lo que conduce a la inhibición del crecimiento de los órganos demandantes y por ende a la baja productividad en muchas especies de plantas (Chen y col., 2018; He y col., 2020).
Martínez y Cayón (2011) indicaron que, en promedio, del área foliar total producida por las plantas de plátano, estas pierden aproximadamente 8 m2 al llegar a la floración y otros 4 m2 adicionales hasta la cosecha; estas pérdidas; sin embargo, podrían ser mayores si no hay un adecuado manejo de las enfermedades foliares o si la planta experimenta condiciones de estrés hídrico. Lo anterior significa que, de la biomasa seca destinada para la formación de hojas durante todo el ciclo, la planta pierde un 60% en forma de hojas secas no recuperables.
3.3 Número de hojas
Por otro lado, no hubo diferencias estadísticas (P>0,05) para el número de hojas emergidas·planta−1 por efecto de los tratamientos evaluados. Los promedios alcanzados para el número de hojas totales (38,33 hojas), el número de hojas al emerger la inflorescencia (12,33 hojas funcionales) y al momento de la cosecha (5,67 hojas), sugiere que la fertilización con magnesio no tuvo influencia sobre esta variable. Estos resultados fueron similares a los obtenidos por Pinchao (2018) quien señaló 5 hojas a la cosecha y 38 a 40 hojas en todo el ciclo del cultivo. Resultados similares fueron reportados por Herrera y Aristizábal (2003) y Jaramillo y Aristizábal (2004) en trabajos con otros cultivares de plátano.
Según Martínez y Cayón (2011) a partir de la emergencia de la inflorescencia, la planta debe mantener un mínimo de 8 hojas para garantizar el llenado del racimo. Aristizábal (2008) establecieron que, si bien el número de hojas funcionales presentes es importante, lo es mucho más la posición de las mismas en la planta, ya que este hecho es el que determina la contribución que cada una de ellas hace al llenado del racimo. A este respecto, observaron que la tasa de emisión foliar en plátano Hondureño Enano y Dominico Hartón tendió a disminuir linealmente a través del tiempo hasta que la planta emitió la inflorescencia.
3.4 Número de manos y de dedos, longitud y perímetro de los dedos, biomasa de los frutos y del racimo y rendimiento·ha−1
Las variables número de manos y de dedos, longitud y perímetro de los dedos, biomasa de los frutos y del racimo y rendimiento·ha−1, todas relacionadas con los componentes del rendimiento, no presentaron diferencias estadísticas (P>0,05) por efecto de las dosis de MgO aplicadas. No obstante, los valores promedios fueron de 5,44 manos·racimo−1; 26,33 dedos exportables·racimo−1; longitud de los dedos de 31,44 cm; perímetro de los dedos de 47,17 mm; biomasa de los dedos de 350 g; biomasa del racimo entre 10,28 y 11,49 kg y rendimiento entre 22,98 y 25,88 t·ha−1.
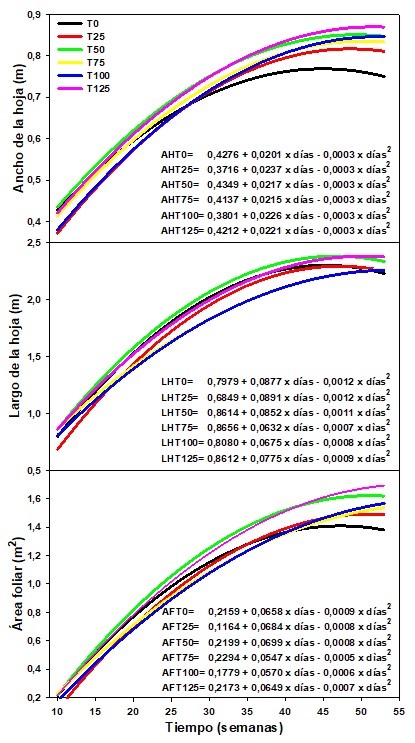
Figura 3. Ancho, largo y área foliar del cultivo de plátano ‘Barraganete’ (Musa ABB), bajo diferentes dosis de fertilización con MgO, en El Carmen, Ecuador. Datos tomados cada 8 días. Se presentan los valores estimados. T0= testigo (0 kg.ha-1 de MgO,negro), T25= 25 kg.ha-1 de MgO (rojo), T50= 50 kg.ha-1 de MgO (verde), T75= 75 kg.ha-1 de MgO (amarillo), T100= 100kg.ha-1 de MgO (azul) y T125= 125 kg.ha-1 de MgO (magenta).
3.5 Eficiencia agronómica (EA) y factor parcial de productividad (FPP)
Las dosis de 50 a 125 kg·ha−1 generaron EA negativas desde −3,33 a −30,67 kg·ha−1, mientras que la dosis de 25 kg·ha−1 fue de 23,95 kg·ha−1 (Figura 4). Las EAs negativas fueron producto de que T0 (testigo sin MgO; 25 277,78 kg·ha−1) tuvo un rendimiento superior a las parcelas fertilizadas, a excepción de T25 (25 876,54).
La intersección de las líneas se considera el punto óptimo, donde el fertilizante fue mayormente aprovechado por la planta para incrementar la producción. La misma fue superior a los 25 kg·ha−1, con una eficiencia agronómica inferior a los 20 kg de fruto·kg·ha−1 de MgO aplicado, lo cual sugiere realizar más investigaciones donde se utilicen dosis más pequeñas, en intervalos entre 20 y 30 kg·ha−1, que permita determinar la dosis idónea para aumentar el rendimiento del cultivo (Figura 4).
Dosis superiores a los 50 kg·ha−1 de MgO disminuyeron la eficiencia agronómica bajo las condiciones de experimentación. Según Avellán y col. (2020) y Cobeña y col. (2020) esto podría haber ocurrido debido a que la absorción de nutrientes, por exceso de fertilización, fue superior al requerido por las plantas, generando un efecto negativo, ya que impidió la absorción de otros nutrientes requeridos en mayor volumen y que podrían haber estado presentes en la solución del suelo. Esto sugiere un desbalance nutricional; en otras palabras, la absorción de una mayor cantidad de MgO y menos de otros nutrientes, especialmente N y P, pudo contribuir a disminuir los rendimientos.
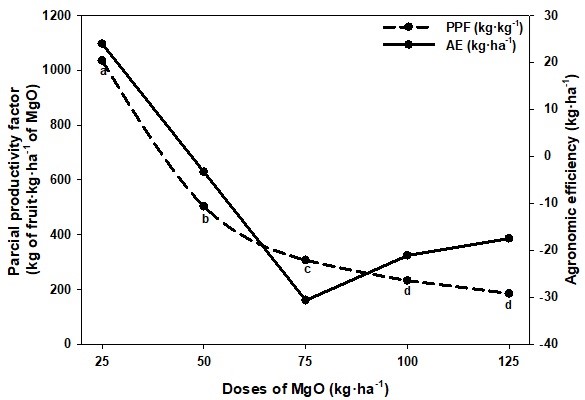
Con relación al FPP, se encontraron diferencias estadísticas (P<0,01) para el FPP por efecto de las dosis de MgO aplicadas. Los valores reportados se encontraron entre 1 035,06 y 184,73 kg de fruto·kg·ha−1 de MgO para las dosis de 25 y 125 kg·ha−1 de MgO, respectivamente (Figura 5). En este sentido, Boaretto, Muraoka y Trevelin (2007) señalaron que en la medida que se incrementó la dosis de fertilizante, la eficiencia productiva de los cultivos fue menor; en otras palabras, con bajas dosis de fertilización la eficiencia del nutriente aplicado fue elevado. Karley y White (2009), y White (2012) indicaron que, en los cultivos perennes como bananos y plátanos, es frecuente la presencia de antagonismos y sinergismos entre nutrientes que afectan el rendimiento, señalando que la relación antagónica entre K, Ca y Mg ha sido la más estudiada, y concluyendo que cuando alguno de estos nutrientes es elevado, disminuyen los contenidos de los otros, generando disminución en el crecimiento y rendimiento de las plantas.
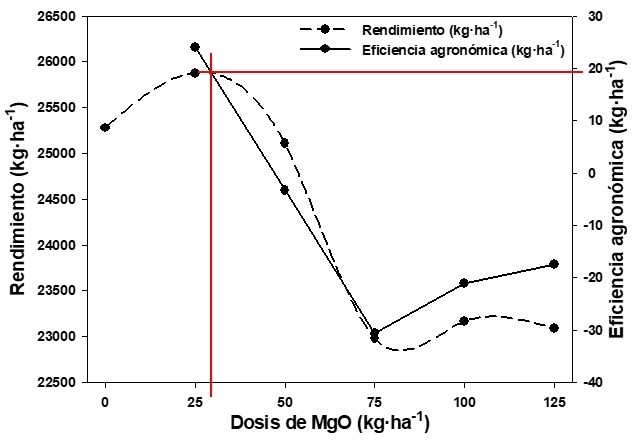
Figura 4. Rendimiento y eficiencia agronómica en plátano ‘Barraganete’ (Musa ABB), bajo diferentes dosis de fertilización con MgO, en El Carmen, Ecuador.
4. Conclusiones
Las variables productivas no fueron influenciadas por las dosis aplicadas, aun cuando el mayor rendimiento, la eficiencia agronómica y el factor parcial de producción se alcanza con la dosis de 25 kg·ha−1 de MgO o cercana a ella, lo cual sugiere realizar investigaciones con dosis cercanas a ese valor. La dinámica de las variables morfo-fisiológicas evaluadas a través del tiempo se ajustan a ecuaciones polinómicas de segundo orden, donde la aplicación de MgO generó diferencias entre las dosis utilizadas en plátano ‘Barraganete’. La dosis de 50 kg·ha−1 de MgO presentó las mayores diferencias entre los tratamientos de fertilización con MgO.

